
-
1 视频
-
2 章节测验
水热和营养环境改变下植物对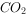 浓度的响应
浓度的响应
植物体含水量一般为60-80%。一方面植物通过根系吸收水分,使地上部分各器官保持一定的膨压,维持正常生理功能;另一方面,植株又通过蒸腾作用散失大量的水分,这一对相互矛盾的过程只有相互协调统一才能保证植株的正常发育。
温度直接影响植物的光合作用、呼吸作用、蒸腾作用,从而影响植物的发芽、开花、结果等生长过程。 植物在一定温度范围内才能正常进行生物化学过程。当环境温度过高或过低,超出生物所能忍受的限度时,生物的生长发育就会受阻,甚至导致死亡。
养分供应状况对植物的生长发育和产量形成有重要的调节作用。
1、与温度的复合作用
一般来讲,随浓度升高,植物光合作用的最适温度会增加5-10℃(Allen, 1994)。
在目前浓度下,C4植物光合作用对温度升高的反应大于C3植物。
2、干旱与浓度升高的复合影响
浓度升高的影响在干早区表现最为明显。大部分实验证明,在水分供应出现紧张时,植物对于
浓度升高表现的正效应更大。
在美国生物圈二号高环境下(平均>2200×
)生长达4年半之久的热带植物与荒漠植物比较,后者对
浓度加倍的反应更明显,如热带植物的净光合作用增加88%,而荒漠植物增加171%;热带植物的水分利用效率增加191%,而荒漠植物增加109%(蒋高明等,1996, 1997) 。
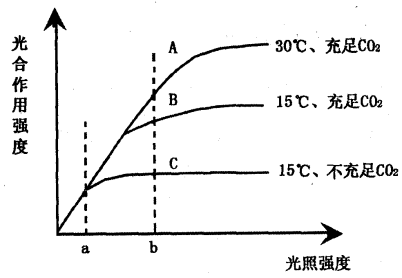
3、不同氮营养水平下植物对的响应
自然生态系统中,氮素营养经常成为生态系统生产力的限制因素。N素供应不足可以通过产物积累,降低光合作用有关酶的表达及限制蛋白质合成导致光合适应现象。

Ziska et al, (1996)发现在播种后34天,高浓度下生长的水稻叶片光合作用高于正常
处理的叶片,与供N水平无关。在52天时,低N水平的叶片光合降低。
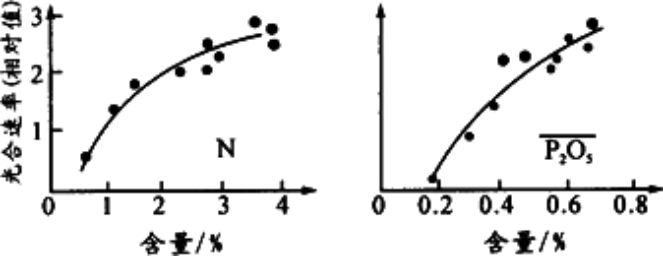
实线表示生物量对N:C的光合反应曲线,虚线为营养限制曲线。A, B分别为在低和高浓度下在同一N:C比的平衡点,C为在高
浓度下满足两种曲线的平衡点。由图可以看出,若在相同的N:C下(即不受营养限制时),高
浓度的促进作用才表现出来。
不同浓度下生物量与叶N含量及N:C比的关系示意图
4、光抑制环境与高浓度下的植物响应
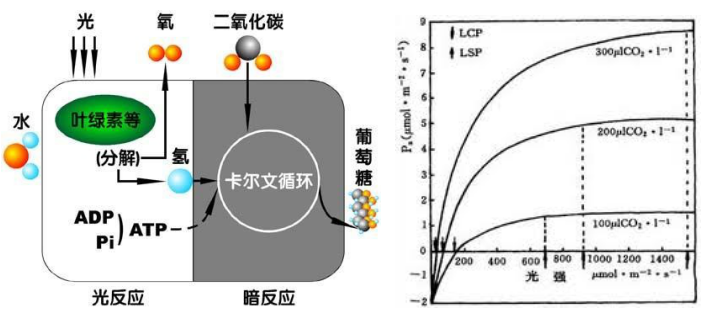
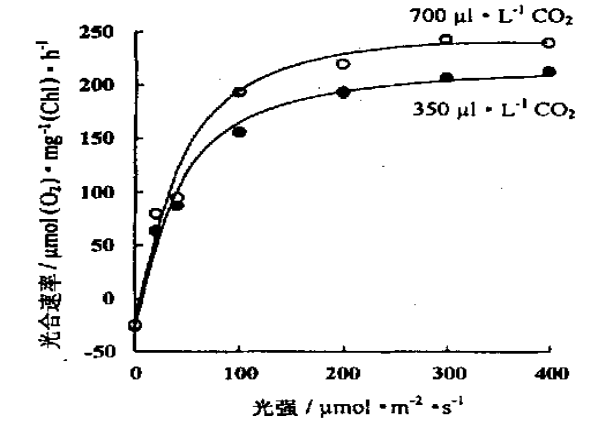
图中是不同浓度培养的斜生栅藻的光合作用—光反应曲线

