
-
1 视频
-
2 章节测验
气候变化
政府间气候变化专业委员会(IPCC WGII, 1995)认为,在未来50或100年间,气候变化对世界渔业的影响可能超过过度捕捞。
鱼是一种变温动物,它们适应温度变化的方法是改变栖息水域,如果其原有栖息水域水温升高,鱼类往往选择向水温较低的更高纬度或外海水域迁移。

全球变暖对中、低纬度鱼群的产量影响较小。
中、低纬度在全球变暖过程中温度变化幅度相对较小
中、低纬度的渔业产量的限制因子主要以营养盐、赤潮和病害为主
相比而言,以光和温度作为主要限制因子的高纬地区的渔业生产受全球变暖的影响要大得多,这也与在全球变暖过程中高纬度水域的水温、风、海流、盐等物理因子变化的幅度较大密切相关。
加拿大、日本、英国和美国的科学家分析了近40年北半球寒温带海水温度与红大马哈鱼栖息范围的动态关系(Williams, 1998),发现未来海洋表层水温变暖的趋势将使极具经济价值的红大马哈鱼从北太平洋的绝大部分水域消失。
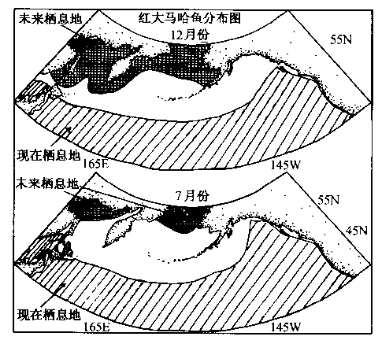
 CO2浓度加倍导致海洋温度上升后红大马哈鱼栖息范围缩小示意图(Willioms, 1998)
CO2浓度加倍导致海洋温度上升后红大马哈鱼栖息范围缩小示意图(Willioms, 1998)
臭氧层破坏
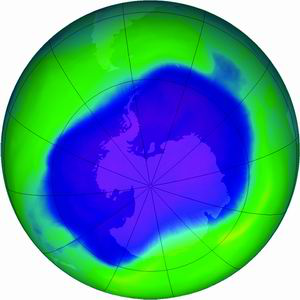
自1986年英国科学家发现南极上空的臭氧空洞以来,其面积有增无减,1996年已扩展至2500万km2。在对氟里昂等直接破坏臭氧层物质的使用加以限制或取缔后,南极臭氧空洞在21世纪初达到最大后可能慢慢缩小。
南极臭氧空洞
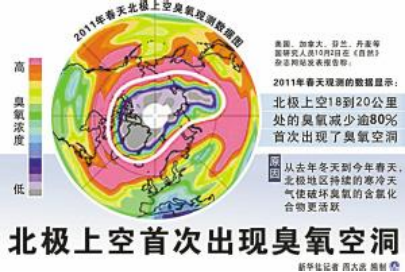
最近,科学家发现北极的臭氧空洞已经形成,并呈不断扩大之势。最令人担忧的是臭氧空洞的形成并不只是与氟里昂和氧化氮等的使用有关。温室气体在加热低空大气的同时将更多的热辐射反射回太空,改变了大气的热传导而使高空平流的大气变冷,也会导致臭氧层受到破坏。
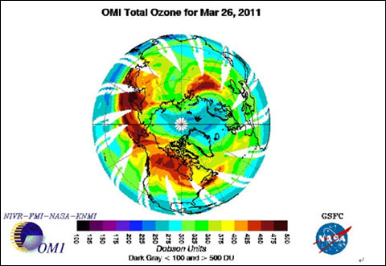
许多研究已经表明紫外辐射对包括浮游植物在内的水生微小生物的生长和繁殖具有损伤作用,导致水域基础生产力下降。
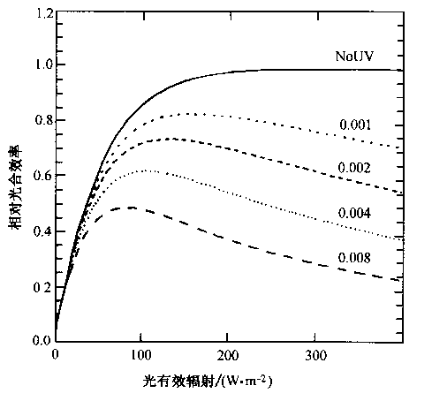
紫外辐射对光合作用的影响,图中数字为实验光照中能引起生物损伤的紫外辐射强度的比例(Cullen, 1992)

