
-
1 视频
-
2 章节测验
磷循环及其与碳、氮循环的耦合作用
硫(sulfur, S)循环
磷(P)是地球上的第10大元素,其原子量为30.975。P在大多数有机体中的重量比仅为1%左右,但它在细胞的能量贮存、传输和利用等方面起着关键作用。
另外,它还制约着生态系统,尤其是水生生态系统的光合生产力,因此,P循环是实现生物圈功能的重要基础。
(1)含磷化合物及其分布
自然界中,磷有4种氧化状态,即-3、0、+3和+5,在这些含磷化合物中,只有以+5价态的磷酸及其化合物在自然界中才是稳定的,他们以各种化合物或以及其可变形态存在于自然界(下表)

植物的光合和呼吸作用控制着磷对生物圈的输入和输出。在陆地生态系统中,几乎所有的生物可利用态磷都是通过含磷矿物的风化产生的。促进风化的因子(如植物根系的活动、根瘤菌的作用)都可提高土壤中的有效磷含量。
一般来说,矿物中的P含量并不高,并且风化的量不足以提供足够多的植物可利用态磷,因此,大多数土壤都或多或少地出现缺磷现象。因此,施用磷肥可显著提高作物的产量。
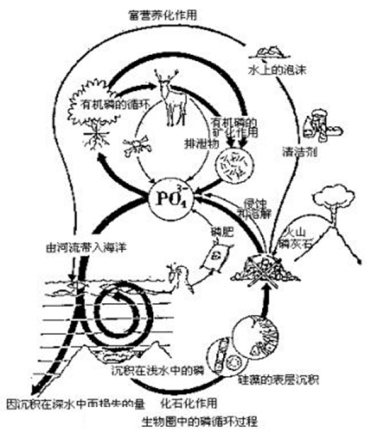
(2)P 循环
全球磷循环与其他几种主要元素循环的一个显著不同之处是几乎没有气体成分参与磷循环。
但90年代由于分析技术的进步,人们已能监测到大气中磷的气态化合物,尽管它在空气中不稳定,平均寿命仅为1d,但毕竞还是有一部分磷经过大气圈进入全球性循环(Glindemann & Bergmann, 1996; Glindemann et al., 1996)。
磷在地球中的含量约为0.1%;
地球总重量为6× Tg;
地球上的P总量为6× Tg;
其中绝大部分P存在于地幔和地核中,只有很少部分的P参与全球的生物地球化学循环。
海洋生物体的P库也可由类似方法求得,即,海洋总生物量为
1800Tg,平均P:C比为1:106,那么,海洋P库则为44 Tg P。表层海洋和深层海洋的P库分别为2800 Tg P和100000 Tg P。
(3)碳、氮和磷循环的耦合作用
在磷循环中,主要通量是由土壤通过径流进入河湖海洋,其中部分颗粒磷沉入海底。由于磷常常是生物生长的限制因素,它的迁移量和库存量会直接影响碳、氮、硫的循环。因此,磷与它们的耦合作用不可忽视(Wollast, 1993)。
磷与碳、氮循环可以在多层次上发生耦合作用。如在分子水平上;细胞水平上;生态系统中,无论是在什么层次的C、N、P元素的迁移, 都是通过生物地球化学循环来发生联系的。
硫(sulfur, S)循环
硫是地壳中的第14大元素,同P相似,S在生物体中的含量也很低,仅为0.25%左右,但对大多数生物的生命过程至关重要。
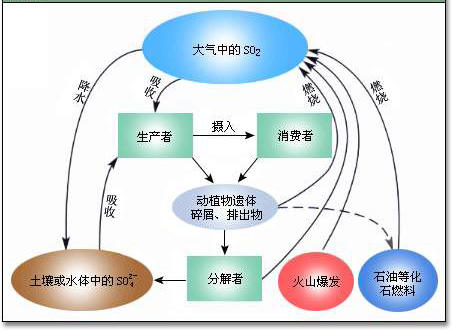
★S在很多自然水体中,存在大量的可溶态,因此,作为养分元素,它很少成为限制因子。
★从全球变化的角度,人们关心S循环是因为它是酸雨和大气气溶胶的主要成分。
★硫的生物地球化学循环研究一直比较活跃,这是由于酸沉降、温室效应乃至臭氧层耗损均与硫的污染有直接或间接的关系。
在自然界中,S可以形成8种氧化状态,其价态从-2到+6,但在地球系统中,常常只发现6种氧状态
碳、氮和硫的生物释放源的未来研究方向
★多样的碳生物释放源
陆地生态系统净生产力的变化以及土地利用的变化这两个不确定因素如何控制碳源的大小
★氮的生物释放源中含氮化合物的系统分析
不局限于常规氮的形态分析,而是系统分析各种无机氮化合物(如,、
、
,
)与有机氮化合物
★硫排放源的重点
重点研究陆地生态系统的源,包括植物与土壤

