-
1
-
2 PPT
我们的机体时刻受到环境中各种病原体的威胁,包括病毒、细菌以及寄生虫等。抵御微生物或其它异物侵入机体的首要屏障是人体的表皮和黏膜层。上呼吸道和肺泡黏膜绒毛的上排运动以及胃肠道动有助于驱逐进入体内的病原体,此外汗液中的溶菌酶对病毒和细菌具有有效的杀伤和抑制作用。然而一旦体表防御机制未能发挥应有的作用,免疫系统与侵入机体的微生物间不可避免地会发生一场“殊死较量”。
一、抗病毒免疫
病毒是地球上除了类病毒以外最简单的生物,由核酸和蛋白质衣売组成,有些病毒还有外膜,其中嵌有膜表面蛋白。病毒体没有独立生存的能力,完全依靠宿主细胞的生物合成系统来复制病毒蛋白及其遗传物质并进行病毒体的组装。
病毒的生命周期分为胞内与胞外两个时期。胞外期的病毒没有任何代谢活动,但具有借助细胞表面受体感染细胞的能力。病毒所利用的细胞表面受体许多与免疫系统有关。例如,人补体受体CR2RNA(CD21)是EB病毒感染B细胞的受体;单纯疱疹病毒能够与CR结合;HIV蛋白通过CD4分子以及趋化因子受体CCR5和CXCR4感染人T细胞和巨噬细胞。与宿主细胞的表面受体结合之后,病毒借助受体介导的吞饮作用进人胞内。胞内的溶酶体酶将包裹病毒核酸物质的蛋白衣壳降解,使病毒的遗传物质暴露于胞浆内并开始指导病毒大分子的合成以及病毒体的组装。少数病毒(如CMV)能够与宿主细胞和平共处,长期寄生,称之为非细胞毒性病毒,多数病毒为细胞毒性病毒,即其在胞内的繁殖和寄生很快导致宿主细胞的死亡。从宿主细胞内获释的病毒体进一步感染更多的邻近细胞,造成局部组织感染灶。病毒也能通过淋巴系统进入血液循环,导致全身感染。病毒对其所感染的宿主细胞有独特的选择性和特异性。有些病毒只能在神经细胞中繁殖,而有些则专门感染肝细胞或者表皮细胞,还有些能够感染全身的多种组织细胞。病毒的这些特点决定了它们所引起疾病的特征。
(一)抗病毒非特异性免疫应答
也叫天然免疫应答,对初次被某种病毒感染的个体来说,由于其尚未建立对该病毒特异的获得性免疫应答,在感染的早期只能依靠天然免疫系统阻止病毒扩散。
1、干扰素(IFN)
是由病毒或IFN诱导剂诱导人或动物细胞产生的一种糖蛋白。
受感染细胞形成并释放IFN,渗入邻近细胞使之产生抗病毒蛋白,抑制病毒蛋白质合成,具有广谱抗病毒、抗肿瘤、免疫调节等作用。
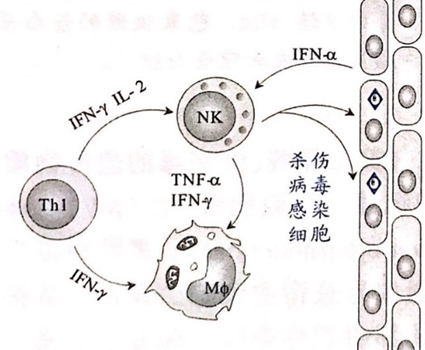
2、NK细胞
通过释放TNF(肿瘤坏死因子)、IFN等物质,可直接杀伤靶细胞(病毒感染细胞,细胞膜发生变化,被NK细胞识别),还可激活巨噬细胞。(如右图)
3、巨噬细胞
巨噬细胞活化后,大量吞噬被补体致敏的病毒体,同时分泌IL-1、IL-6、IL-8、IL-12、TNF-α等细胞因子和趋化因子,促进毛细血管内皮细胞上调黏附分子,以便于中性粒细胞和单核细胞浸润感染局部。
4、补体。
干扰素、NK细胞、巨噬细胞和补体的共同作用使得侵入体内的病毒不能迅速扩散。
(二)抗病毒特异性免疫应答

1、细胞免疫应答
局部组织发生病毒感染时巨噬细胞和NK细胞被活化并分泌TNF-α。TNF-α使组织中的树突状细胞(DC)成熟并携带病毒抗原向引流淋巴结迁移(如右图A),DC在迁移的过程中将病毒抗原加工处理并开始表达大量MHC分子和共刺激分子。一些病毒体和抗原也能够通过淋巴管进入引流淋巴结,被淋巴结内巨噬细胞或者FDC所捕获。在引流淋巴结的皮质区,巨噬细胞以及来自病毒感染局部的DC通过MHC-Ⅰ和Ⅱ类分子将病毒抗原肽递呈于膜表面,活化抗原特异的CD4+T细胞、CD8+T细胞和B细胞,并使其迅速扩增(如右图B)。所产生的Th和CTL效应细胞离开引流淋巴结经淋巴循环系统抵达被感染局部(如右图C)。
由于病毒蛋白是在胞浆内复制的,病毒抗原肽可以被MHC-Ⅰ分子递呈。而表达于细胞表面的病毒体或者蛋白经内吞后进入MHC-Ⅱ抗原递呈途径。因此,病毒感染通常能够诱导较强的CTL和Th细胞反应。宿主初次感染3-4天之后外周开始出现病毒特异的CTL和Th细胞,7-10天时达到高峰。局部组织中的APC通过递呈病毒抗原肽激发CTL和Th细胞发挥免疫效应。Th细胞分泌更多的细胞因子,加强免疫应答。CTL迅速消灭被病毒感染的细胞,消灭病毒赖以藏身之处和复制“工厂”。
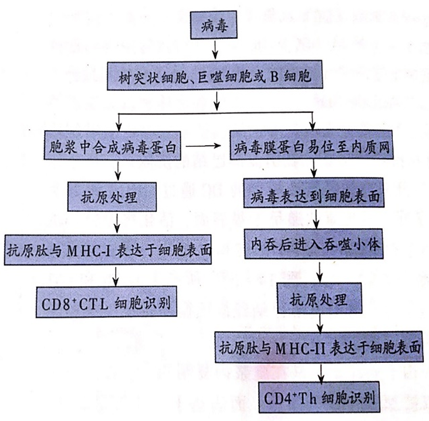
2、体液免疫应答
初级淋巴滤泡中的FDC捕获随淋巴液而来的病毒体或者病毒蛋白,并将其展示于细胞表面。在病毒特异性Th细胞的帮助下,能够识别病毒抗原的B细胞被活化并在淋巴滤泡中进行多次分裂。所产生的B细胞分化为浆细胞,分泌针对病毒的IgM、IgG、IgA抗体。B细胞同时通过Ig基本超突变进一步微调其BCR,产生与病毒抗原具有高亲和力的抗体。
抗体在抗病毒感染中具有十分重要的作用(如下图)。首先,抗体与病毒表面蛋白质的结合可以阻断其与细胞表面受体的接触或者干扰病毒向细胞内的侵入,黏膜表面的IgA分子在这方面的作用尤为重要。其次,抗体分子结合病毒后通过激活补体导致病毒体的裂解,在此过程中产生的C3a和C5a等补体片段可趋化白细胞。抗体还能够有效地致敏病毒颗粒,使其更容易被吞噬细胞所识别和吞噬。有些病毒膜蛋白能够表达于宿主细胞的表面。病毒特异性抗体通过对这些病毒蛋白的识别而“标记”被感染细胞,诱发补体或者NK和巨噬细胞对这些细胞的杀伤。
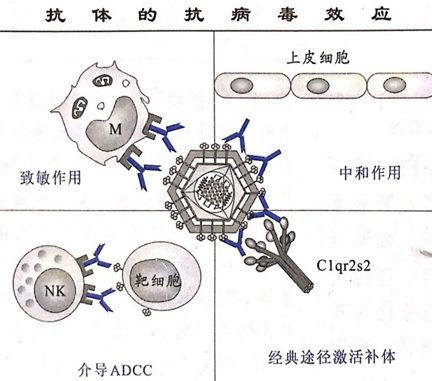
三、抗细菌免疫应答
1、抗细菌非特异性免疫应答
穿过体表或者黏膜层的细菌首先可能激活补体而被裂解(下图A)。补体活化后产生的C3a和C5a片段参与激活肥大细胞并趋化其它炎症细胞(下图B)。此外,细菌及其产生的毒素对神经未稍、肥大细胞和巨噬细胞均构成强烈的刺激,有可能直接导致肥大细胞脱颗粒,启动局部的炎症反应(下图C)。在各种趋化介质的作用下,血液循环中的中性粒细胞和单核细胞浸润感染局部,吞噬并消化入侵的细菌(下图D),非特异性免疫应答在细菌感染的早期控制细菌的快速增殖,有时在特异性免疫应答启动之前就已经将入侵的病原微生物清涂,但多数情况下需要特异性免疫系统的参与才能实现这一目标。
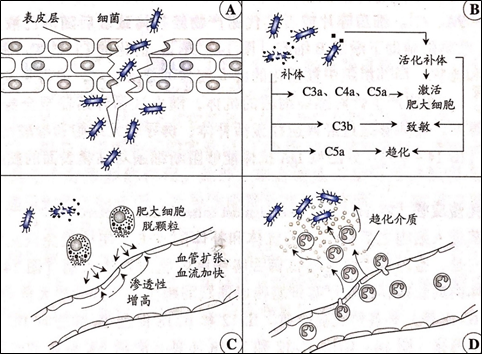
2、抗胞外寄生菌免疫应答
胞外寄生菌对机体的危害主要来自其所产生的各种外毒素和内毒素,这类细菌所诱导的特异性免疫应答的主要效应是①通过抗体中和细菌毒素并调理(致敏)病原菌;②促进吞噬细胞对细菌的识别、吞噬和消化(如下图)。
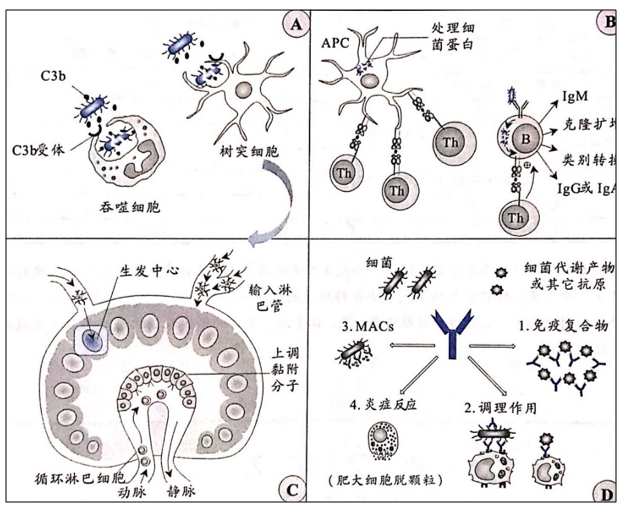
组织中的DC吞噬细菌碎片或者抗原后迁入引流淋巴结并在那里激活T细胞(上图A、B、C)。细菌碎片或者其代谢产物经补体致敏后随淋巴液进入引流淋巴结,进而在Th细胞的辅助下激活B细胞(上图B)。活化的CD4+Th细胞随血液循环进入外周组织和器官。局部组织中被活化的肥大细胞可分泌IL-4,使得免疫应答向Th2偏移。B细胞产生针对感染细菌的抗体,随血液循环运输至全身。特异性抗体可以中和细菌毒素,还能够激活补体、诱导肥大细胞和嗜酸性粒细胞的活化与脱颗粒反应(上图D)。分泌型IgA抗体能够阻断细菌对黏膜表面的黏附。
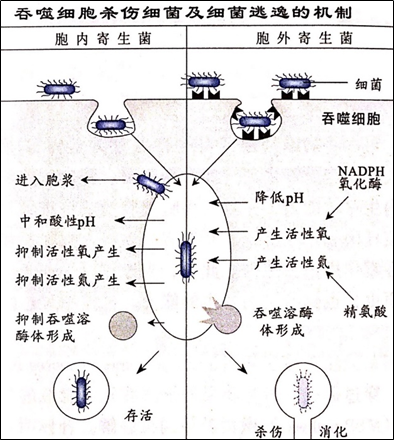
3、抗胞内寄生菌免疫应答
胞内寄生菌进入细胞之后能够躲避抗体和补体的攻击,使体液免疫应答难以发挥效应。更为重要的是,胞内寄生菌能够抵御吞噬细胞的胞内杀伤机制(如下图)。在胞浆或者吞噬体内的长期存在对巨细胞构成强烈的刺激,使其分泌大量的细胞因子(如IL-1、IL-12和IL-18等)和各种炎症介质。L-12和L-18促进Th细胞向Th1分化,使胞免疫应答向Th1偏移。L-12和L-18还具有激活NK细胞的作用。Th1和NK细胞通过它们分泌的IFN-γ进一步激活巨噬细胞,使其进入亢奋状态,三者之间形成正反馈激活环路,加大免疫应答的强度。因此,针对胞内生寄生菌,尤其是分枝杆菌(如结核杆菌)的免疫应答往往导致较为严重的Ⅳ型超敏反应,甚至可以形成慢性肉芽肿结节。应该指出,所有的病原微生物感染均同时诱导体液和细胞免疫应答,只不过不同病原微生物所诱导反应的侧重不同。