
第二节 基 因
一、基因的概念及种类
(一)基因的概念
1865年遗传学奠基人孟德尔通过8年豌豆杂交试验提出生物的各种性状是由细胞内的遗传因子决定的。1909年遗传学家约翰森首次用“gene”(基因)这个名词来表示遗传因子,并沿用至今。而后经过100多年的发展,特别是20世纪90年代以来,随着分子生物学和分子遗传学的迅猛发展,人类对基因结构和功能的认识日新月异。目前普遍认为基因是具有特定遗传效应的DNA片段,是遗传功能的基本单位,即基因是能够表达基因产物的完整DNA片段。
(二)基因的种类
根据人类体细胞基因组DNA的碱基排列顺序重复程度的不同,可把基因组DNA分为单一序列、重复序列、多基因家族和拟基因。
1.单一序列 单一序列(unique sequence)是指单拷贝的DNA序列,在一个基因组中只出现一次或几次。10kb以下的单一序列较多,而大于25kb的单一序列较少。它们主要是构成细胞内编码蛋白质或酶的基因,称为结构基因(structure gene)。人类体细胞基因组中含有2.5万~3.5万个结构基因,其中一部分是分散存在的,一部分是形成多拷贝的基因家族存在的,它们常常被多拷贝的重复序列的DNA片段所隔开。单一序列占人类基因组中DNA总量的60%~65%。
2.重复序列 重复序列(repetitive sequence)是指这种DNA序列在基因组中拷贝几十份、几百份或几十万份,占人类基因组中DNA总量的30%~35%。它们大多没有编码功能,它们可维持染色体的结构和稳定、参与细胞分裂或具有调控作用。其详细的生物功能有待进一步的研究。根据DNA片段长度和拷贝数,可分为高度重复序列和中度重复序列。
(1)高度重复序列:在人类基因组中高度重复序列(highly repetitive sequence)的碱基序列通常较短,长度小于200bp,碱基重复次数为106~108,约占人类基因组中DNA总量的10%。
(2)中度重复序列:中度重复序列(moderately repetitive sequence)的碱基序列较长,长度大于200bp,重复次数为102~105,在人类基因组中是分散存在或成簇存在的,占人类基因组中DNA总量的20%~30%。
3.多基因家族 多基因家族(multigene family)是指由一个祖先基因经过重复和变异所形成的一组基因,它们在基因组中拷贝数有微小差异,功能相同或相关,分为两类:一类是一个基因多次拷贝,序列几乎相同,成簇存在于一条染色体上,它们同时表达或在不同时间表达,表达的基因产物相同,功能相同;另一类是一个多基因家族中不同成员成簇分布于几条不同的染色体上,这些基因序列虽有不同,基因表达产物也不同,但功能相关。例如:编码红细胞上血红蛋白-珠蛋白的基因就是一个多基因家族,它由α珠蛋白基因簇和β珠蛋白基因簇组成;α珠蛋白基因簇成簇存在于16号染色体短臂上,β珠蛋白基因簇成簇存在于11号染色体短臂上;两种珠蛋白基因簇序列高度一致,来自同一祖先珠蛋白基因,属于一个多基因家族。
4.拟基因 拟基因(pseudogene)又称为假基因,是指在多基因家族中,有些基因和有功能的基因结构相同或相近,但不能表达基因产物的基因,如组蛋白、干扰素、α珠蛋白、β珠蛋白、肌动蛋白质以及人的tRNA和rRNA的多基因家族中均有拟基因存在。拟基因可能是有功能的基因由于缺失、倒位、点突变等原因而失活,变成了无功能的基因。拟基因通常是由有功能的基因通过几次突变生成的,大多数的多基因家族中都有拟基因,但是在基因组中所占比例很小。
二、基因的结构
生物基因的DNA序列包括编码序列(编码区)和侧翼序列两部分。原核生物的基因是一个连续编码的DNA分子片段,即编码区全部具有编码功能。真核生物包括人类基因,其编码区是不连续编码的,一些具有编码功能的DNA序列被一些非编码DNA序列隔开,形成镶嵌排列的断裂形式,称为断裂基因,如图3-4所示。
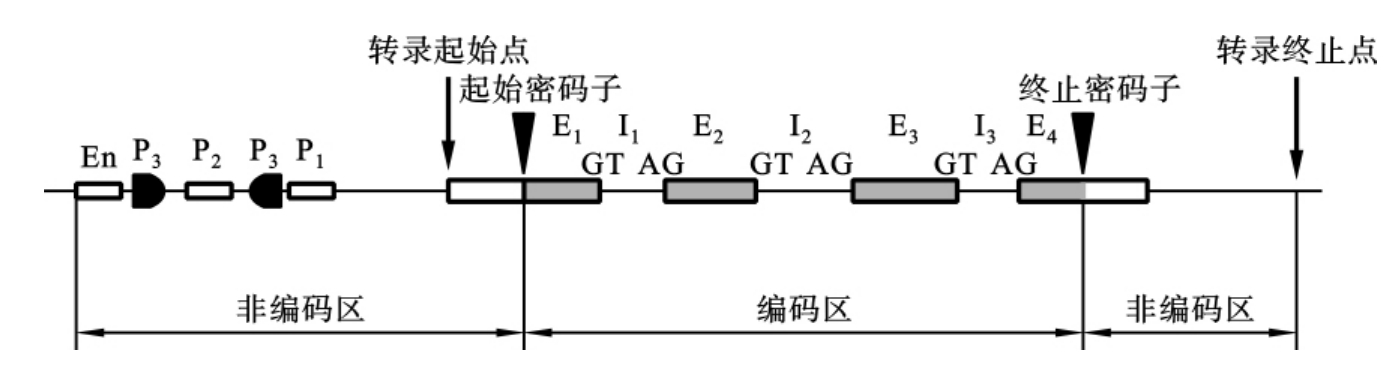
图3-4 真核生物结构基因的结构示意图
注:En为增强子,P为启动子(P1为TATA框,P2为CAAT框,P3为GC框),E为外显子,I为内含子。
(一)内含子和外显子
在真核生物的断裂基因中,编码序列被非编码序列所分隔。其中具有编码功能的DNA序列,称为外显子(exon,E)。它是基因中可表达为多肽的部分。两个外显子之间的非编码DNA序列,称为内含子(intron,I)或插入序列(insertion sequence,IVS)。内含子的大小和数目是不相同的,如人的假肥大性进行性肌营养不良(DMD)基因有75个外显子和74个内含子,全长2 300kb,编码3 685个氨基酸。真核生物断裂基因中的内含子在转录后被切除,然后把外显子连接在一起,再经过一系列的加工,才能成为有生物活性的成熟的mRNA,经细胞核的核孔复合体被运送到细胞质中,在核糖体上翻译出特定的基因产物。真核生物断裂基因的外显子和内含子的接头区是高度保守的一致序列,称为外显子-内含子接头,即在每个内含子的5′端开始的两个核苷酸为GT,3′端末尾的两个核苷酸为AG,这种接头方式称为“GT-AG法则”。这两组碱基是真核细胞基因中普遍存在的,这种特殊的碱基序列是hnRNA剪切加工成为成熟的mRNA的信号,正因为有这种特殊的碱基序列,真核生物遗传物质表达的初始产物hnRNA中没有编码功能的内含子才能在酶的催化作用下被准确地切除。
在人类基因组中,也有少数的结构基因无内含子的序列,如位于Y染色体上的SRY基因等,其编码序列呈连续状态,无内含子。同时真核生物断裂基因中的内含子和外显子的关系并不是固定不变的,有时同一条DNA链上的某一段DNA序列,当它作为编码一条多肽链的基因时是内含子,而作为编码另一条多肽链的基因时是外显子,结果是同一基因的DNA序列可以转录为两种或两种以上的mRNA。
(二)侧翼序列
真核生物基因结构中的第一个外显子和最后一个外显子的外侧存在的非编码区,称为侧翼序列(flanking sequence)。在侧翼序列中有一些对基因表达具有调控作用的DNA序列,称为调控序列(regulator sequence)。调控序列不被转录,但对基因的有效转录具有调控作用,又称为顺式作用元件,它包括启动子、增强子和终止子等。
1.启动子 启动子(promoter,P)是指与转录启动有关的特定序列,是位于基因转录起始点上游的100~200bp范围内的一段特异的核苷酸序列,是RNA聚合酶的结合部位,能促进转录过程。它包括以下三种结构序列。
(1)TATA框:位于转录起始点上游-27~-19bp处,是高度保守的一段DNA序列,其序列由TATAATTAT7个碱基组成,为富含GC的DNA序列。TATA框通过与转录因子TFⅡ结合,再与RNA聚合酶结合形成复合物,能够精确识别转录起始点,并控制转录的水平。
(2)CAAT框:位于基因转录起始点上游-80~-70bp范围内的一段特异的核苷酸序列,其序列由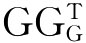 CAATCT 9个碱基组成,该序列只有一个碱基可以变化。CAAT框通过与转录因子CTF结合,控制转录的启动和频率。
CAATCT 9个碱基组成,该序列只有一个碱基可以变化。CAAT框通过与转录因子CTF结合,控制转录的启动和频率。
(3)GC框:位于CAAT框的两侧,有两个拷贝,其序列为GGCGGG,GC框通过与转录因子Sp1结合,起增强转录效率的作用。
并非所有真核生物的启动子都同时含有上述三种结构框序列。
2.增强子 增强子(enhancer)是指一段能增强启动子转录效率的特定序列,通常为存在于启动子上游或下游3kb处或更远处的一段核苷酸序列,本身不具有启动子活性,但它可以增强启动子的转录能力,提高基因的转录效率。不同基因的增强子碱基序列差异较大,但有一个核心序列为GGTGTG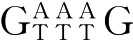 ,该序列有三个碱基可以变化。增强子发挥作用无明显方向性,可以是5′-3′方向,也可以是3′-5′方向。这是由于DNA可以折成环状使增强子与启动子结合,或使增强子与RNA聚合酶结合形成复合物。例如,人类的β珠蛋白基因的增强子是由两个相同序列的长度为72bp的核苷酸序列串联重复组成的,存在于转录起始点上游约-1 400bp处,或存在于转录起始点下游约-3 300bp处,可以使转录效率提高200倍。
,该序列有三个碱基可以变化。增强子发挥作用无明显方向性,可以是5′-3′方向,也可以是3′-5′方向。这是由于DNA可以折成环状使增强子与启动子结合,或使增强子与RNA聚合酶结合形成复合物。例如,人类的β珠蛋白基因的增强子是由两个相同序列的长度为72bp的核苷酸序列串联重复组成的,存在于转录起始点上游约-1 400bp处,或存在于转录起始点下游约-3 300bp处,可以使转录效率提高200倍。
3.终止子 终止子(terminator)是一段具有终止功能的特定DNA序列,是位于基因3′非编码区下游的一段反向重复的碱基序列,是一段由5′AATAAA3′组成的回文序列,为转录终止信号。AATAAA是多聚腺苷酸(polyA)的附加信号,也是RNA聚合酶停止加工的信号,该序列转录后可以形成发卡式结构,可以阻止RNA聚合酶的移动。由于该发卡式结构末尾的一串U与DNA模板中的A结合不稳定,从而使转录产物RNA从模板上脱落下来,使转录终止。因此终止子的终止作用不在自身DNA序列上,而在于它的转录产物RNA。
三、基因的功能
遗传物质的主要功能是复制遗传信息,从而使亲代的遗传信息准确地传给子代,表达遗传信息,从而使生物体的生命现象得以体现,这就是基因的复制与表达。
(一)遗传信息的储存
遗传信息是以DNA链上4种碱基(A、G、C、T)的不同组合形式而储存的,特定的碱基序列包含了特定的遗传信息。这种遗传信息可通过转录传递到mRNA分子上,作为蛋白质合成的指令,指导蛋白质的生物合成。
(二)基因的复制
基因的复制(replication)是指以亲代DNA分子的每一条链为模板,利用游离的三磷酸脱氧核苷酸(dNTP)为原料,在酶的催化作用下,在碱基互补配对原则的指导下合成子代DNA分子的过程。基因的复制是以DNA的复制为基础的,通过复制和细胞分裂,使亲代的遗传信息准确地传给子代,所以DNA复制在保持物种延续、遗传的稳定性方面发挥着重要作用。
1.自我复制 在原核生物的双链闭合环状裸露DNA分子中,DNA的复制往往只有一个复制起始点,从复制起始点开始双向复制,在复制中形成一个Q形的分子后,两个复制叉交于一点而形成两个双链闭合环状裸露DNA子代分子。高等真核生物的双链线形DNA分子的复制起始点有多个,从每个复制起始点开始双向复制,当DNA分子上的复制起始点全部连通后,DNA分子的复制就完成了,如图3-5所示。含有一个复制起始点的复制单位称为复制子(replicon)。复制子仅有起点而无终点。每个复制子大小为30~300bp,人类的一个基因组大约含有105个复制子。真核生物种类不同,复制子大小也不同,同一生物在不同生理条件下,复制子的大小也不同,个体生长快时复制子小。
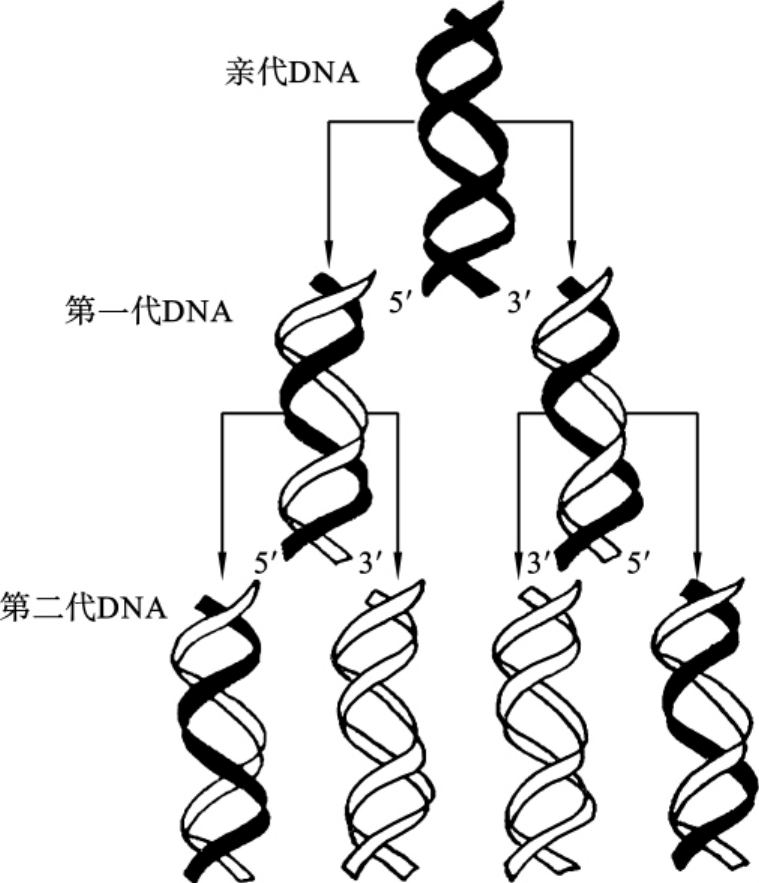
图3-5 DNA分子自我复制
由于DNA分子特殊的双链互补结构使得复制形成的两个子代DNA分子和亲代DNA分子的结构和组成完全相同,而且新合成的子代DNA分子都保留了一条作为模板的母链(亲代DNA),因此DNA分子的自我复制又称为半保留复制(semiconservative replication)。DNA分子的半保留复制保证了生物体前、后代之间遗传性状的相对稳定。
2.半不连续复制 DNA分子的复制是双向复制,每一条亲代DNA分子链都可以作为模板,但是由于DNA聚合酶只能使脱氧核苷酸连在多聚脱氧核苷酸链游离的3′端碳位的羟基上,所以新合成的DNA子链只能沿5′-3′方向进行。以3′-5′方向的DNA母链为模板,互补合成的5′-3′的子链符合DNA聚合酶的特性,先合成一段RNA作为引物(primer),以提供游离脱氧核苷酸连接所需的3′碳位的羟基,然后游离脱氧核苷酸一个一个连接上去,使复制形成的DNA子链以5′-3′方向延伸,复制是连续的,复制速度较快,所用时间短,称为前导链(leading strand)。而以5′-3′方向的DNA母链为模板,互补合成的5′-3′的子链不符合DNA聚合酶的特性,复制是不连续的,其DNA分子的复制是当作为模板的DNA分子解旋到一定位置时先合成一段RNA作为引物,以提供游离脱氧核苷酸连接所需的3′碳位的羟基,然后游离脱氧核苷酸一个一个连接上去,使复制形成的DNA子链是带有引物RNA的以5′-3′方向延伸的DNA片段。我们把这段带有引物RNA的DNA片段称为冈崎片段(Okazaki fragment)。由于以5′-3′方向的DNA母链为模板,互补合成的DNA子链不符合DNA聚合酶的特性,复制是不连续的,必须先合成一段一段带有引物RNA的DNA片段,然后把引物RNA切去,把缺口补起来,再把DNA片段连起来,所以复制速度较慢,所用时间长,称为后随链(lagging strand)。由此看来DNA分子的复制是半不连续复制(semi-discontinuou replication),如图3-6所示。
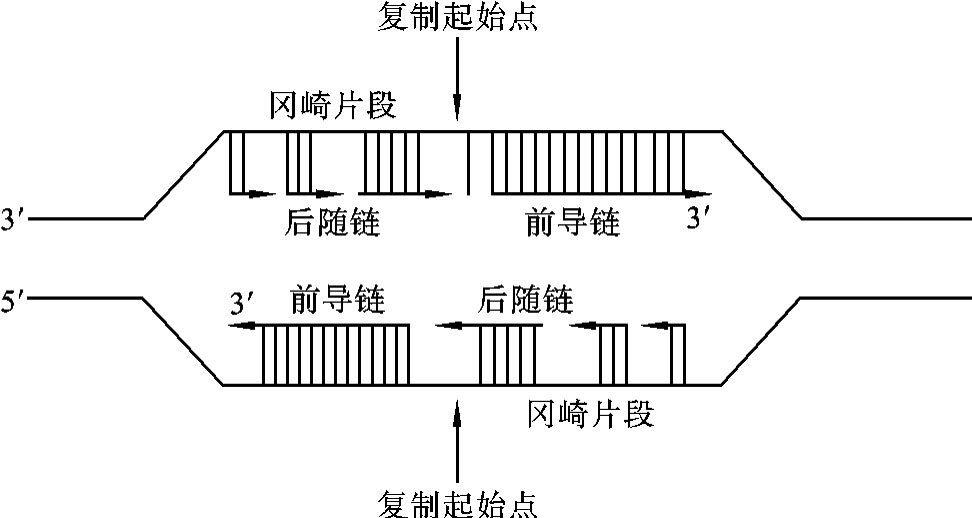
图3-6 基因的半不连续复制
3.DNA分子的复制需要RNA引物 由前所述的DNA分子的复制过程可以看出,DNA聚合酶只能使脱氧核苷酸连在多聚核苷酸链游离的3′端碳位的羟基上,它不能在复制时直接起始DNA新链或冈崎片段的合成,因此,在DNA复制时,必须首先在RNA聚合酶的作用下,以亲代DNA为模板,先合成一段RNA作为引物,这段RNA称为引物RNA。
4.DNA分子复制的方向是5′-3′方向 新合成的子代DNA链是由5′端向3′端方向延伸的,这是由DNA聚合酶的特性决定的。
(三)基因的表达
基因的表达(expression)是指基因的DNA分子所储存的遗传信息转化为蛋白质分子的氨基酸的种类、数目和排列顺序的过程。它包括转录和翻译两个过程。基因控制蛋白质合成的过程是:以DNA为模板,合成RNA,然后以mRNA为模板合成蛋白质,如图3-7所示。原核生物没有被核膜包裹的成形的细胞核,故而转录和翻译是在同时间和同地点同步进行的,而且转录形成的mRNA直接与核糖体结合指导蛋白质的合成。真核生物有被核膜包裹的成形的细胞核,转录是在细胞核内进行的,而翻译是在细胞质中进行的。
1.转录 转录(transcription)是指以基因DNA分子的一条特定链为模板,利用游离的三磷酸核苷酸(NTP)为原料,在酶的催化作用下,在碱基互补配对原则的指导下合成RNA分子的过程。
(1)转录过程 在基因DNA分子的两条脱氧核苷酸链中,只有一条链是储存遗传信息的,这条脱氧核苷酸链称为编码链,而另一条与它互补的链称为反编码链,转录是以基因DNA分子的反编码链为模板,经过转录形成的RNA分子的碱基排列顺序和编码链的碱基排列顺序一致,只是DNA分子中的碱基“T”在RNA分子中被碱基“U”所取代了。这样经过转录把DNA分子所储存的遗传信息转化为RNA分子的碱基排列顺序,将来通过RNA带到细胞质中的核糖体上指导蛋白质的合成,从而使基因的DNA分子所储存的遗传信息转化成为蛋白质分子的氨基酸的种类、数目和排列顺序。RNA的合成方向为5′-3′方向,如图3-8所示。
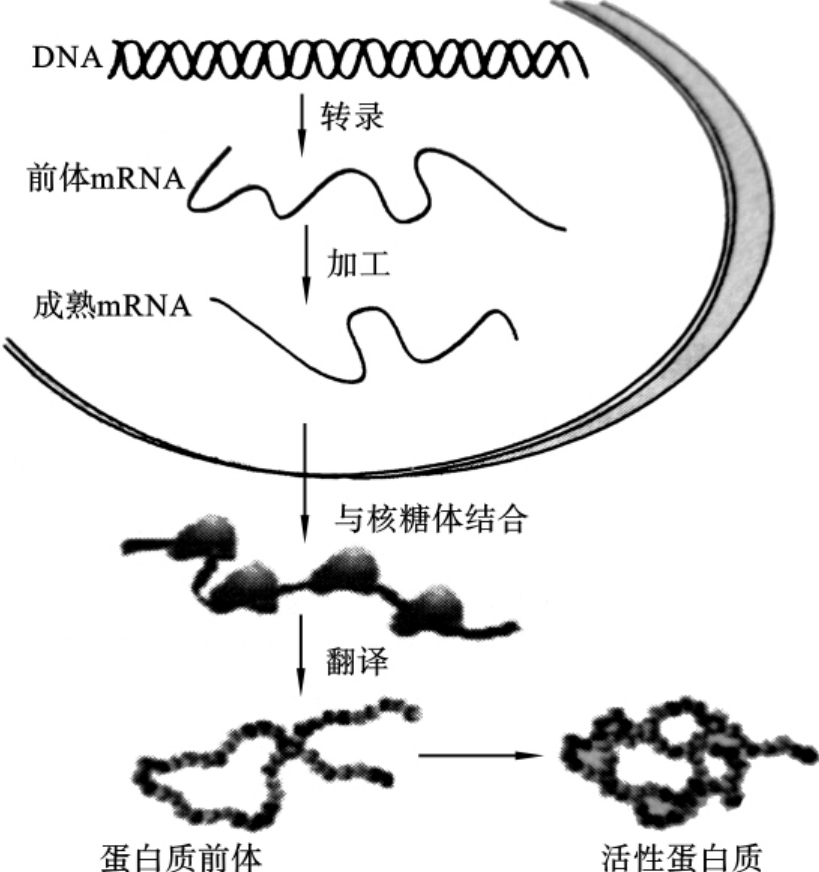
图3-7 真核生物的基因表达过程
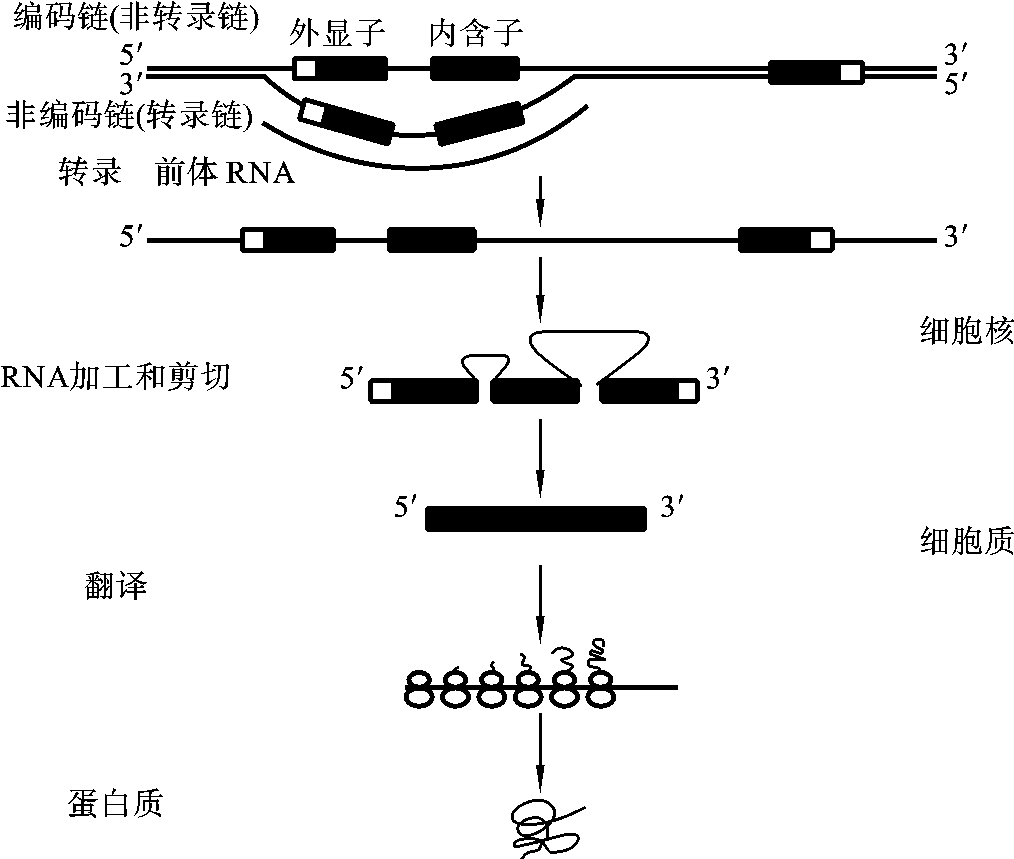
图3-8 基因表达示意图
DNA的转录需要RNA聚合酶的催化作用,RNA聚合酶有三种,分别为RNA聚合酶Ⅰ、RNA聚合酶Ⅱ和RNA聚合酶Ⅲ。酶不同产物也不一样,转录产物分别是tRNA、mRNA和rRNA。
(2)转录产物的加工和修饰 原核生物基因的编码区是连续的,都具有编码功能,转录形成的mRNA可以直接指导蛋白质的合成。而真核生物的断裂基因的编码区是不连续的,由外显子和内含子组成,在转录时把外显子、内含子和部分的侧翼序列都包含了,转录形成hnRNA。hnRNA要经过进一步加工才能成为成熟的有生物活性的mRNA,指导蛋白质的合成。hnRNA的加工一般包括剪接、戴帽和添尾等过程。
①剪接:剪接(splicing)是指在酶的催化作用下,将hnRNA中的内含子切去,然后把相邻外显子按顺序准确拼接成为连续编码的mRNA的过程。剪接过程严格遵循“GU-AG”原则,即剪接点是由基因的内含子与外显子交接处的剪接信号(5′GTAG3′)决定的,在RNA其剪接信号就是5′GU-AG3′。
在hnRNA的剪接过程中细胞核内的小RNA(small nuclear RNA,snRNA)起着很重要的作用。snRNA是存在于真核生物中的一段高度保守的富含尿嘧啶(U)的RNA,长约250bp,又称为U族RNA,现已发现的有U1、U2、U3、U4、U5、U6等。这些snRNA与一些蛋白质结合形成snRNP(small nuclear ribonucleoprotein)。snRNP通过RNA-RNA互补,可以识别内含子中的RNA特定序列,并与内含子序列结合形成snRNP-内含子复合物,使内含子的5′GU和3′AG聚合在一起,内含子形成一个套状结构。通过两次转酯反应,相邻的外显子由酯键相连,使内含子的5′GU和3′AG结合并由酯键相连,snRNP-内含子复合物从hnRNA分子上脱落下来,此时snRNP与内含子分离,再去识别内含子中的RNA特定序列。在整个hnRNA的剪接过程中,各种snRNP形成剪接体,协助hnRNA的剪接。
②戴帽:戴帽(capping)是指在hnRNA加工成为成熟的有生物活性的mRNA的过程中,在RNA的5′端的第一个核苷酸连接一个甲基化的帽,即7-甲基鸟苷酸帽(m7′G)。戴帽的作用可以有效封闭mRNA的5′末端使其不再加核苷酸,同时也可以保护5′末端,使其不受磷酸酶和核酸酶的水解作用,以达到增强mRNA稳定性的作用。此外,戴帽也可以促使mRNA到达细胞质中与核糖体的小亚基识别并结合,促进翻译的进行。
③添尾:添尾(tailing)是指在hnRNA加工成为成熟的有生物活性的mRNA的过程中,在RNA的5′端戴帽的同时,在其3′端连接100~200个腺苷酸,形成多聚腺苷酸(polyA)尾。这一过程需要腺苷酸聚合酶的催化作用,称为多聚腺苷酸化反应。
添尾的作用目前尚不完全清楚,其作用可能是稳定mRNA的3′端,使其免受酶的催化作用,同时可以促使mRNA由细胞核运送到细胞质中。
经以上加工形成的有生物活性的mRNA,进入细胞质中的核糖体上作为模板,指导蛋白质合成的过程就是翻译。
2.翻译 翻译(translation)是指转录形成的mRNA的碱基排列顺序所储存的遗传信息在细胞质中的核糖体上“解读”成为蛋白质分子中的氨基酸的种类、数目和排列顺序的过程,其实质是以mRNA为模板指导合成蛋白质多肽链的过程。
翻译是在细胞质中的核糖体上进行的,有许多RNA参与蛋白质的合成过程,其中mRNA是蛋白质合成的模板,tRNA在蛋白质合成过程中作为转运器,它能特异性地与活化的氨基酸结合并形成氨酰基tRNA,从而不断地把氨基酸运送到细胞质中的核糖体上,参与蛋白质的合成。每一种氨基酸只能由一种或几种特定结构的tRNA来负责转运。rRNA是蛋白质合成场所——核糖体的组成成分。
(1)遗传密码 遗传密码(genetic code)是指转录形成的成熟的mRNA的碱基排列顺序所储存的遗传信息,将来mRNA到达细胞质中的核糖体上指导蛋白质合成时它要决定氨基酸到底放到什么位置。经研究发现mRNA核苷酸链上相邻的三个核苷酸序列对应一个氨基酸,称为三联体密码或密码子(codon),密码子是遗传密码的单位。对于组成mRNA的四种碱基A、U、G、C,如果每三个碱基对应一个氨基酸,将有43=64,即有64种组合形式,也就是有64个密码子。在64个密码子中,有三个密码子,即UAA、UAG、UGA不编码任何氨基酸,是多肽链合成的终止信号,称为终止密码子,剩余61个密码子具有编码氨基酸的功能,遗传密码表见表3-2。
表3-2 遗传密码表
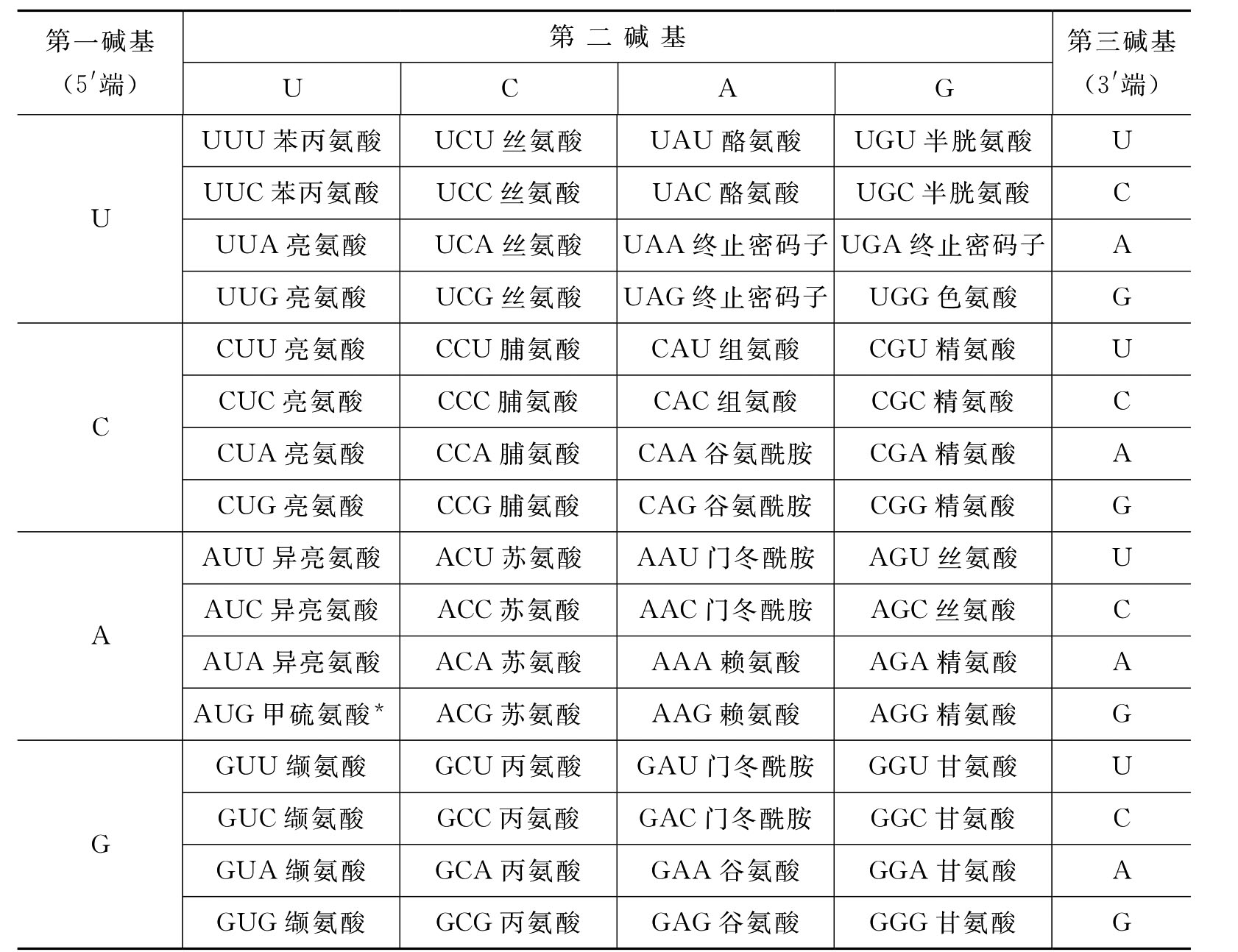
*AUG在原核生物中为甲酰甲硫氨酸,在真核生物mRNA分子5′端起始部位是起始密码子,同时编码甲硫氨酸,在其他部位编码甲硫氨酸。
遗传密码具有以下一些特性。
①通用性:遗传密码在生物界是从病毒、细菌、真菌到高等生物和人类都通用的,称为遗传密码的通用性,但是也有少数例外,如AUG在真核生物细胞是编码甲硫氨酸(蛋氨酸),而在原核生物中则是编码甲酰甲硫氨酸,此外,在线粒体中,UGA是编码色氨酸的,而不是终止密码子,CUA是编码苏氨酸的,而不是编码亮氨酸的,AUA是编码甲硫氨酸的,而不是编码异亮氨酸的。
②兼并性:组成蛋白质分子的氨基酸有20种,而密码子有61种,这就决定一种氨基酸的密码子不一定是一种,大多数氨基酸都有两种以上的密码子。我们把多个密码子对应同一种氨基酸的现象称为遗传密码的兼并性,如ACU、ACC、ACA、ACG共同编码苏氨酸。
③方向性:遗传密码是从mRNA的5′端开始向3′端一个一个阅读,不重叠,无标点。
④摇摆性:tRNA在运送氨基酸时,是靠tRNA的反密码环上3个相邻的碱基-反密码子与mRNA上的密码子碱基互补配对,从而保证tRNA准确地运送氨基酸,但是由于遗传密码具有兼并性,同一tRNA运输同一氨基酸到达具有兼并性的不同密码子时,密码子与反密码子的碱基互补配对的第一位、第二位的碱基是严格遵循碱基互补配对原则的,而第三位的碱基就比较灵活了,可以不配对,这称为遗传密码的摇摆性。
此外,在遗传密码表中,AUG如果位于mRNA的5′端的起始处时,它就是起始密码子,是蛋白质合成的起始信号,在其他位置时则是编码甲硫氨酸(蛋氨酸)或甲酰甲硫氨酸的密码子。
(2)翻译过程 ①起始:翻译开始时,在起始因子(initiation factor,IF)的作用下,核糖体的30S小亚基结合到mRNA分子的起始密码子AUG部位,形成“30SmRNA-IF”复合体;然后,甲酰蛋氨酰-tRNA进入复合体,tRNA分子上的反密码子与mRNA分子上的起始密码子AUG互补结合;最后,核糖体的50S大亚基与30S小亚基结合形成70S起始复合体,起始因子释放。②延长:核糖体上有两个氨酰-tRNA的结合位点,即供体部位(P位)和受体部位(A位),在70S起始复合体中,甲酰蛋氨酰-tRNA结合在P位,第二个氨酰-tRNA结合在A位,在转肽酶催化下,P位上的氨酰-tRNA所携带的氨酰基移至A位,与A位上的氨基酰脱水缩合形成二肽,空载的tRNA离开P位,在移位酶和GTP供能的情况下,核糖体沿mRNA的5′-3′方向移动1个密码子,即A位上的氨酰-tRNA移位到P位,空载的A位又可接受1个新的氨酰-tRNA,不断重复此过程,肽链不断增长,所合成的多肽链中,氨基酸的种类、数目及排列顺序完全由mRNA上的密码子决定。③终止:当mRNA分子上出现终止密码子(UAA、UAG、UGA)时,核糖体停止移动,肽链停止延长,在释放因子的参与下,多肽链从核糖体上释放,同时,核糖体的大亚基和小亚基分离,mRNA与核糖体分离,如图3-9所示。
蛋白质合成速度很快,大肠杆菌合成1个含300~400个氨基酸的蛋白质仅需要10~20s。通常1个mRNA分子可以同时结合多个核糖体,即同一mRNA可依次翻译出多条相同的多肽链。在整个翻译过程中一个mRNA分子通常与多个核糖体结合,最多可达60多个,形成多聚核糖体(polyribosome)。这样多个核糖体以同一mRNA分子作模板,同时进行翻译,按不同进度形成多条相同的多肽链,从而大大提高了翻译的进程。
(3)遗传信息表达的中心法则 遗传物质所储存的遗传信息通过复制而使亲代的遗传信息准确地传给子代,通过转录和翻译的表达过程从而使遗传信息在生物体的生命现象中得以体现。复制、转录、翻译便构成了遗传信息的流向,称为信息流(information flow)。
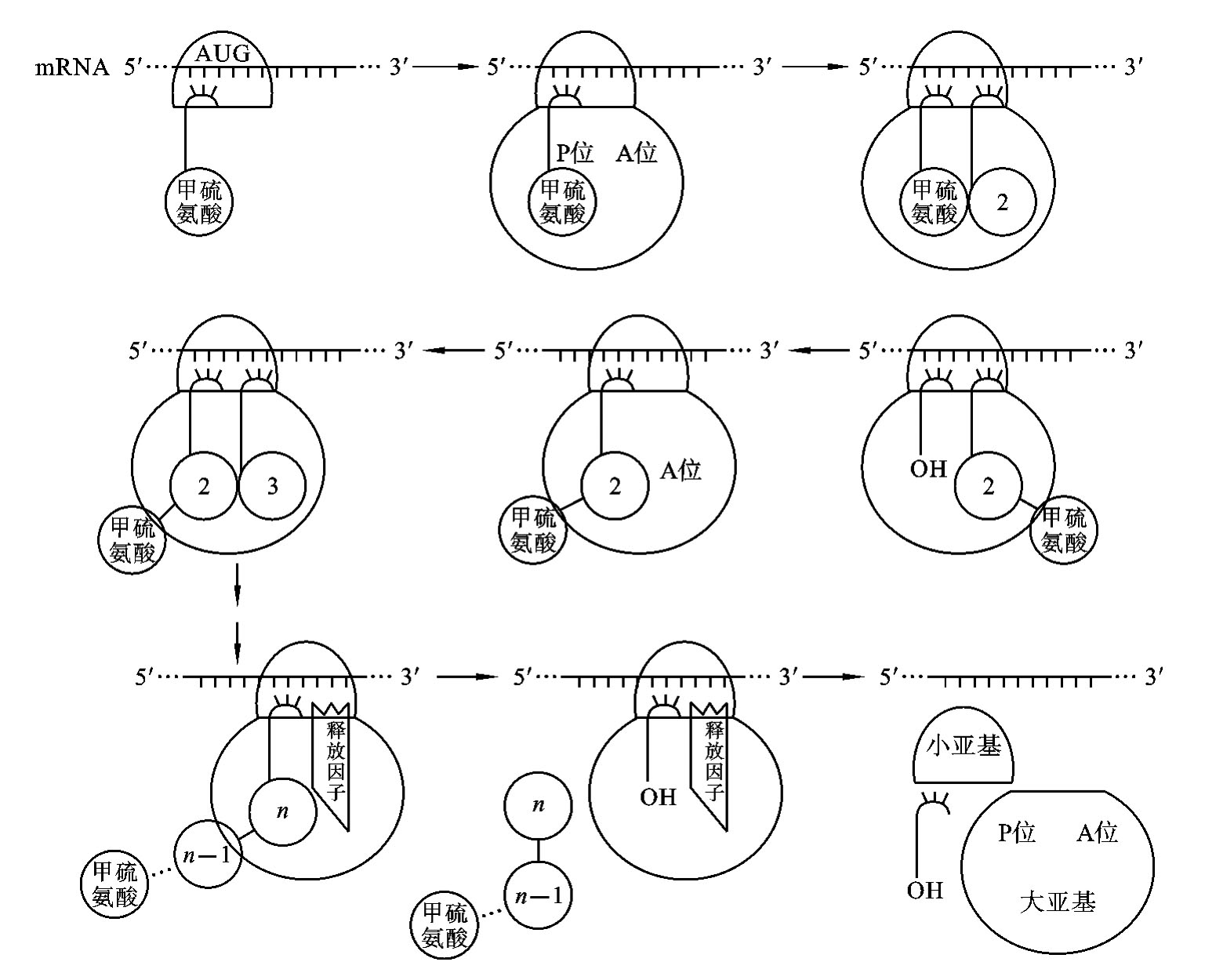
图3-9 蛋白质合成过程示意图
1958年克里克提出,信息流的方向只能为DNA→RNA→蛋白质,是单向不可逆传递的,遗传信息不可能由蛋白质传给蛋白质,也不可能由蛋白质传给DNA或RNA。克里克把这种信息传递的原则称为中心法则(central dogma)。1970年巴尔的摩和特明等发现了RNA病毒的逆转录现象,以及一些单链RNA病毒的自我复制和翻译,对中心法则进行了补充和完善,如图3-10所示。
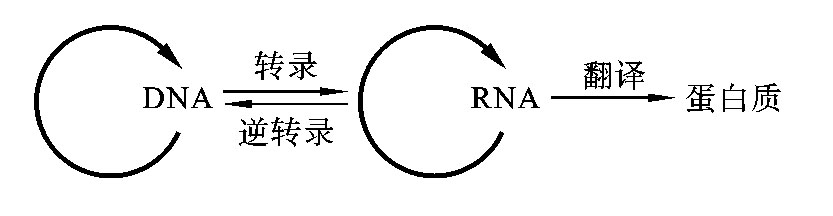
图3-10 中心法则图解
(四)基因表达的调控
1.原核生物的基因表达调控 原核生物基因表达的调控主要在转录水平上,多以操纵子为单位进行,操纵子既是基因转录单位,也是基因的调控单位。20世纪60年代初,法国分子生物学家雅各布和莫若最早在大肠杆菌中发现大肠杆菌对于乳糖代谢的调节是由乳糖操纵子(lactose operon)控制的。乳糖操纵子只在环境中有乳糖存在时才开始合成相关酶系,乳糖分解完之后,酶的合成也就停止了,从而调节和控制细胞的发育。
2.真核生物的基因表达调控 真核生物的每个体细胞中有两个基因组。每个基因组在一定时期内、在一定条件下只有部分特定的基因在表达,这表明存在着有效的基因调控系统。由于真核生物的细胞结构比较复杂,遗传物质主要在被完整核膜包被的细胞核内转录,而翻译发生在细胞质内,因此遗传信息在细胞内有一个转移过程,所有过程都需在适当的调节下进行,才能使遗传物质得以表达,使生物体出现一定的表型,所以真核生物的基因调控比原核生物要复杂得多,它包括转录前、转录中、转录后、翻译、翻译后五个水平的调控。因此,真核生物基因表达的调控比原核生物的要复杂得多,它是在不同层次、不同水平上进行的。

