第三章 种群及其基本特征
第三、四、五章是种群篇,通过种群基本特征、生活史、种内与种间关系的学习,认识种群是生态学各层次中最重要的一个层次,又是群落结构与功能的最基本单位,许多与环境变化相联系的生物变化都发生在这一层次。 生物资源的开发与保护都涉及种群的基本知识和基本理论,在实施生态恢复、生态工程等生态建设方面,具有重要意义。
第一节 种群的概念与动态
一、种群的概念
种群(population)是占有一定空间的同种个体的组合。 除生态学外,进化论、遗传学、分类学和生物地理学等也都使用种群这个术语。 由于学科不同,为了强调各自的侧重面,有的学者还在种群定义中加进诸如能相互进行杂交、具有一定结构、一定遗传特性等内容。(种群是一定时间内占有一定空间的同种个体的组成或集合)
在应用种群概念时,既可是抽象的,也可是具体的。 在了解种群的基本特征和基本理论时均用抽象概念,当落实到某个具体的种群研究时,如某块森林中的梅花鹿种群,则要根据代表性限定其空间,乃至时间范围,这个范围的大小是由研究者划分的。 例如大至全世界的蓝鲸,小至一块草地上的克氏针茅。 实验室饲养的一群小家鼠,也可称为一个实验种群。
从生态学观点来看,种群是生物群落和生态系统的基本组成单位,是种间关系的基本单位,是生物资源开发利用的具体对象。
二、种群的基本特征
自然种群有3个基本特征:
数量特征——个体的生生死死使种群大小不断变化。
空间特征——种群不仅有分布区域,也有在生活空间中的布局。
遗传特征——种群内个体可相互交配,具有一定的基因组成,系一个基因库,以区别于其他物种。
当然,地理空间跨度较大的广布种,不同种群会发生一定的地理变异,使该物种的遗传多样性增加。如果环境在地理空间上连续变化,则导致种群基因频率或表型的渐变,表型特征或等位基因频率逐渐改变的种群叫做渐变群(cline)。
种群生态学是研究种群动态、特征及其生态规律的科学,如种群的数量、分布、生活更格局以及种群与其栖息环境中的非生物因素和其他生物种群的相互关系等。 此外,与种群生态学有密切关系的种群遗传学研究种群的遗传过程,包括选择、基因流、突变和遗传漂变等。 20世纪60年代,很多生物学家认识到分别研究种群生态学和种群遗传学均具有局限性,发觉种群中个体数量动态和个体遗传特性动态有密切的关系,因此,力图将这两个独立的分支学科有机地整合起来,生态遗传学和进化生态学就是在这种思想影响下迅速发展起来的,从而丰富了种群生物学的内容。
三、种群的动态
种群动态是种群生态学的核心内容。种群动态研究种群数量在时间上和空间上的变动规律,包括:1种群的数量或密度; 2种群的分布;3种群数量变动和扩散迁移;4种群调节。
1.种群的密度
严格说来,密度(density)和数量(number)是有区别的,在生态学中应用数量高、数量低、种群大或种群小这些概念时,有时虽然没有指明其面积或空间单位,但也必然将之隐含在其中。 因为没有空间单位的数量多少是无意义的。
研究种群动态规律,首先要进行种群的数量统计(census)。进行统计前,首先要确定被研究种群的边界。因为许多生物种呈大面积的连续分布,其边界是模糊的,所以在实际工作中往往随研究者的方便来确定种群边界。 在数量统计中,种群大小最常用的指标是密度。 密度通常以单位面积(或空间)上的个体数目表示,对于病虫害等微生物和小型动物,也有用每片叶子、每个植株、每个宿主为单位的。 最直接的方法是计数种群中每一个个体,如一片林子中所有的某个树种、繁殖基地上所有的海豹等。用航空摄像可计数所有移动中的羚羊,或间隔较远的大型仙人掌。这种数量调查适用于有限范围内的种群。
样方法是最常用的统计方法,即在若干样方中计数全部个体,然后以平均数来估计种群整体。样方必需有代表性,通过随机取样来保证结果的可靠性,并用数理统计方法来估计变差和显著性。
由于生物的多样性,具体数量统计方法随生物种类或栖息地条件而异;大体分为绝对密度和相对密度两类。绝对密度是指单位面积或空间的实有个体数,而相对密度则只能获得表示数量高低的相对指标。 植物和动物的计算方法不同。在调查和分析种群密度时,首先应区别单体生物和构件生物。
(1)单体生物:单体生物(unitary organism)的个体很清楚,如蛙有四条腿,昆虫六条腿等,各个个体保持基本一致的形态结构,它们都由一个受精卵发育而成。大多数动物属单体生物。
单体生物的数量估计方法有多种形式,例如10只/hm2 黄鼠是绝对密度,而每置100铗,日捕获10只是相对密度,即10%捕获率。相对密度又可分为直接指标和间接指标,如10%捕获率是表示黄鼠只数的直接指标,而每hm 鼠洞数则是间接指标。
对不断移动位置的动物,直接统计个体数很困难,可以应用标志重捕法。在调查样地上,捕获一部分个体进行标志后释放,经一定期限进行重捕。根据重捕取样中标志比例与样地总数中标志比例相等的假定,来估计样地中被调查动物的总数,即
M:N=m:n
式中:N——样地上个体总数;M——标志数;n——重捕个体数;m——重捕中标志数。
对于许多动物,由于获得绝对密度比较困难,相对密度指标成为有用资料,诸如捕获率,遇见率,洞口、粪堆等活动痕迹,鸣声,毛皮收购量,单位渔捞的渔获量等。
(2)构件生物:构件生物(modular organism)由一个合子发育成一套构件(modules)组成的个体,如一株树有许多树枝,一个稻丛有许多分蘖,不仅构件数很不相同,而且从构件还可产生新的构件,其多少随环境条件而变化。 高等植物是构件生物,营固着生活的珊瑚、薮枝虫、苔藓等也是构件生物。
如果说对于单体生物以个体数就能反映种群天小,那么对于构件生物就必须进行两个层次的数量统计,即从合子产生的个体数(它与单体生物的个体数相当)和组成每个个体的构件数。 只有同时有这两个层次的数量及其变化,才能掌握构件生物的种群动态。
对于许多构件生物,研究构件的数量与分布状况往往比个体数(由合子发展起来的遗传单位)更为重要。 一丛稻可以只有一根主茎到几百个分蘖,个体的大小相差悬殊,所以计算秆数比区分主茎更有实际意义。 乔木可能有若干个水平的构件,果树上的枝节还具有不同年龄,有叶枝和果枝的区别,每一果座上花数与果实数也有变化。 许多天然植物都有无性繁殖,个体本身就是一个无性系的“种群”。 由此可见,研究植物种群动态,必须重视个体水平以下构件组成的“种群”的重要意义,这是植物种群与动物种群的重要区别。
2.种群统计学
种群具有个体所不具备的各种特征,这些特征多为统计指标。
①初级种群参数,包括出生率(natality)、死亡率(mortality)、迁入和迁出。出生和迁入是使种群增加的因素,死亡和迁出是使种群减少的因素。出生率是一广义的术语,泛指任何生物产生新个体的能力,不论这些新个体是通过分裂出芽、结籽、生产等哪一方式产生。迁出是指种群内个体由于种种原因而离开原种群的领地,迁入则是别的种群进人领地。
②次级种群参数,包括性比(sex ratio)、年龄结构(age structure)和种联增长率等。种群统计学就是种群的出生、死亡、迁移、性比和年龄结构等的统计学,最初用于人口统计上,现用于一切生物。
(1)种群年龄结构和性比:研究种群的年龄结构和性比对深入分析种群动态和进行预测预报具有重要价值。种群的年龄结构是指不同年龄组的个体在种群内的比例和配置情况。用不同宽度的積柱从上到下配置而成的图称年龄锥体(age pyramid)。横枉的高低位置表示由幼到老的不同年龄组,宽度表示各年龄组的个体数或百分比。按锥体形状,年龄锥体可划分为3个基本类型(图3-1)。

图3-1 年龄锥体的3种基本类型(Kormondy,1996)
(a)增长型种群;(b)稳定型种群;(c)下降型种群。
①增长型种群:锥体呈典型金字塔形,基部宽,顶部狭,表示种群有大量幼体,而老年个体较少。种群出生率大于死亡率,是迅速增长的种群。
②稳定型种群:锥体形状介于(a)、(c)两类之间,老、中、幼比例大体相同。出生率与死亡率大致相平衡,种群稳定。
③下降型种群:锥体基部比较狭、而顶部比较宽。 种群中幼体比例减少而老体比例增大,种群死亡率大于出生率,是不断衰退的种群。
性比是种群中雄性个体和雌性个体数目的比例,哺乳动物受精卵的♂/♀比大致是50:50,这叫做第一性比,幼体成长到性成熟这段时间里,由于种种原因,♂/♀比还要继续变化,到个体成熟时为止,这时♂/♀的比例叫做第二性比,以后还会有充分成熟的个体性比,叫做第三性比。 性比对种群的配偶关系及繁殖潜力有很大影响。在野生种群中,因性比的变化会发生配偶关系及交配行为的变化,这是种群自然调节的方式之一。
(2)生命表的编制:生命表(life table)是一种有用的工具。简单的生命表只是根据各年龄组的存活或死亡数据编制,综合生命表则包括出生数据,从而能估计种群的增长。
Conell(1970)对某岛上1959年出生并固着在岩石上的所有藤壶(Balanus glandula)进行逐年的存活观察,其结果见表3-1中的x和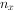 栏,到第9年全部死光。生命表有若干栏,其含义:x为按年龄的分段,为x期开始时的存活数,
栏,到第9年全部死光。生命表有若干栏,其含义:x为按年龄的分段,为x期开始时的存活数,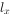 为x期开始时的存活数,
为x期开始时的存活数,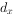 为从x到x+1的死亡数,
为从x到x+1的死亡数,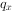 为从x到x+1的死亡率,
为从x到x+1的死亡率,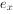 为x期开始时的生命期望或平均余年。
为x期开始时的生命期望或平均余年。
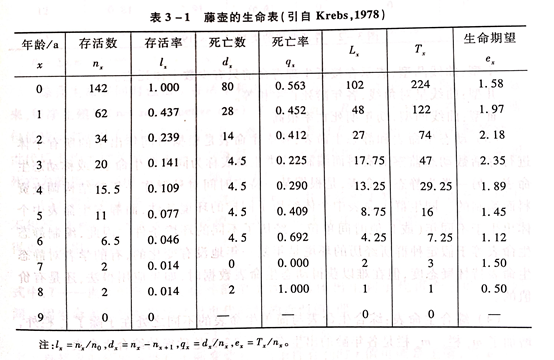
 和
和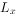 栏一般不列入表中,但为计算方便,需要和栏。是从x到x+1期的平均存活数,即=(+
栏一般不列入表中,但为计算方便,需要和栏。是从x到x+1期的平均存活数,即=(+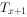 )/2。则是进人x龄期的全部个体在进入x期以后的存活个体总年数,即
)/2。则是进人x龄期的全部个体在进入x期以后的存活个体总年数,即 。例如:
。例如: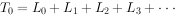 ,
, 。
。
从这个生命表可获得3方面信息:
①存活曲线(survivorship curve),以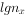 栏对x栏作图可得存活曲线。 存活曲线直观地表达了该同生群(cohort)的存活过程。Deevey(1947)曾将存活曲线分为3个类型(图3-2);藤壶的存活曲线接近I型。
栏对x栏作图可得存活曲线。 存活曲线直观地表达了该同生群(cohort)的存活过程。Deevey(1947)曾将存活曲线分为3个类型(图3-2);藤壶的存活曲线接近I型。
②死亡率曲线,以栏对x栏作图。 藤壶在第一年死亡率很高,以后逐渐降低,接近老死时死亡率迅速上升。
③生命期望,表示该年龄期开始时的平均能存活的年限。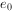 为种群的平均寿命,藤壶
为种群的平均寿命,藤壶 。
。

图3-2 存活曲线的类型(Krebs,1985)
Ⅰ型:曲线凸型,表示在接近生理寿命前只有少数个体死亡。
Ⅱ型:曲线呈对角线,各年龄死亡率相等。
Ⅲ型:曲线凹型,幼年期死亡率很高。
(3)动态生命表和静态生命表:藤壶生命表是根据对同年出生的所有个体进行存活数幼态监察的资料而编制,这类生命表称为同生群生命表,或称动态生命表。 另一类为静态生命表,是根据某一特定时间对种群作一年龄结构调查资料而编制的。 同生群生命表中个体经历了同样的环境条件,而静态生命表中个体出生于不同年(或其他时间单位),经历了不同的环境条件。 因此,编制静态生命表等于假定种群所经历的环境是年复一年地没有变化的,有的学者对静态生命表持怀疑态度,但在难以获得动态生命表数据时,如果应用得法,还是有价值的。
(4)综合生命表:综合生命表与简单生命表的不同之处在于除了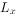 栏外,增加了
栏外,增加了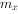 栏。栏是各年龄的出生率。表3-2是猕猴的综合生命表。
栏。栏是各年龄的出生率。表3-2是猕猴的综合生命表。
表3-2 南湾猕猴雌猴生命综合表(引自江海声等,1989)
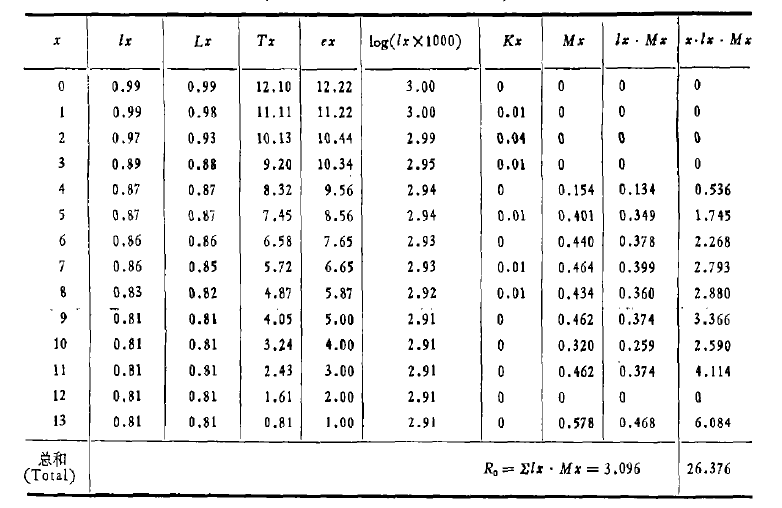
综合生命表同时包括了存活率和出生率两方面数据,将两者相乘,并累加起来,得净生殖率(net reproductive rate):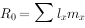 ,
,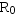 代表该种群(在生命表所包括特定时间中的世代净增值率猕猴生命表的
代表该种群(在生命表所包括特定时间中的世代净增值率猕猴生命表的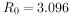 ,表示猕猴数经一世代后平均地增长到原来的3.096倍。 该生命表还增加
,表示猕猴数经一世代后平均地增长到原来的3.096倍。 该生命表还增加 栏,是表示年龄组死亡力的指标,它由栏导出,即
栏,是表示年龄组死亡力的指标,它由栏导出,即 。
。
(5)种群增长率(r)和内禀增长率(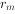 ):由于各种生物的平均世代时间并不相等,作种间比较时,世代净增殖率(
):由于各种生物的平均世代时间并不相等,作种间比较时,世代净增殖率(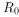 )的可比性并不强,种群增长率r值则显得更有应用价值。r值可按下式计算:
)的可比性并不强,种群增长率r值则显得更有应用价值。r值可按下式计算:
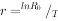
式中:T——世代时间,它是指种群中子代从母体出生到子代再产子的平均时间。用生命表资料可以估计出世代时间的近似值,即:
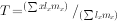
自然界的环境条件在不断变化着,当条件有利时r值可能是正值,当条件不利时可能变为负值。 因此,长期观察某种群动态时,自然种群增长率r值的变化是很有用的指标。 但是,为了进行比较,在实验室条件下,人们能排除不利的天气条件,提供理想的食物,排除捕食者和疾病,在这种人为的“不受限制”的条件下,就能观察到种群的内禀增长率(innate rate of increase)。 例如林昌善等(1964)曾测定杂拟谷盗实验种群的值,=0.074/d,即该种群以平均每日每雌增加0.074个雌体而增长。
从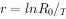 式来看,r值随R。增大而增大,随T值增大而变小。 例如,计划生育的目的是要使r值变小,据此式有两条途径:①降低R。值,即使世代增殖率降低,这就要限制每对夫妇的子女数;②使T值增大,即可以通过推迟首次生殖时间或晚婚来达到。在计划生育中,后一种途径是不应忍视的。Cole曾用模型证明:假如妇女生育年龄从15岁推到25岁,则人口增长率就会有相当大的降低。
式来看,r值随R。增大而增大,随T值增大而变小。 例如,计划生育的目的是要使r值变小,据此式有两条途径:①降低R。值,即使世代增殖率降低,这就要限制每对夫妇的子女数;②使T值增大,即可以通过推迟首次生殖时间或晚婚来达到。在计划生育中,后一种途径是不应忍视的。Cole曾用模型证明:假如妇女生育年龄从15岁推到25岁,则人口增长率就会有相当大的降低。
总之,生命表的实质是描述种群生死过程的一种有用图表模式。由于生物的生活史五花八门,生命表的形式也随之有很大的区别。在今日植物种群生态学和动物种群生态学发展趋向于统一的形势下,一些著名的种群生态学家已存按生活史类型划分生命表上迈开一大步。
3.种群的增长模型
种群动态模型研究在近几十年来有较大发展,它是理论生态学的主要内容,对种群生态学做出了重要贡献。 数学模型是用来描述现实系统或其性质的一个抽象的、简化的数学结构。 科学工作者用它来揭开此系统的内在机制和对系统行为进行预测。建立动植物种群动态数学模型的目的,是阐明自然种群动态的规律及其调节机制毅助理解各种生物和菲生物因素是怎样影响种群动态的。
模型研究必须从客观实际出发来建模,最后又必须通过实践来检验。 在数学模型研究中,人们最感兴趣的不是特定公式的数学细节,而是模型的结构及其参数的生物学或生态学意义及解释,即哪些因素决定种群的大小,哪些参数决定种群对自然和人为干扰的反应速度等。
(1)与密度无关的种群增长模型:种群在“无限”的环境中,即假定环境中空间、食物等资源是无限的,因而其增长率不随种群本身的密度而变化,这类增长通常呈(指数式增长)可称为与密度无关的增长(density-independent growth,或译为非密度制约性增长)。
与密度无关的增长又可分为两类,如果种群的各个世代彼此不相重叠,如一年生植物和许多一年生殖一次的昆虫,其种群增长是不连续的、分步的,称为离散增长,一般用差分方程描述;如果种群的各个世代彼此重叠,例如人和多数兽类,其种群增长是连续的,用微分方程描述。
①种群离散增长模型 最简单的单种种群增长的数学模型,通常是把世代t+1的种群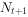 与世代t的种群
与世代t的种群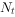 联系起来的差分方程:
联系起来的差分方程:
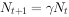 或
或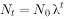
式中:N——种群大小;t一一时间;λ一一种群的周限增长率。
例如,一年生生物(即世代间隔为一年)种群,开始时10个雌体,到第二年成为200个,那就是说,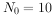 ,
,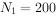 ,即一年增长到20倍。今以λ代表两个世代的比率:
,即一年增长到20倍。今以λ代表两个世代的比率:

如果种群在无限环境下以这个速率年复一年地增长,即10,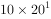 ,
,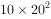 ,
,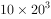 ,...,
,...,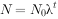 。
。
将方程式两侧取对数,即:

它具有直线方程y=a+bx的形式。因此,以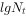 对t作图(图3-3a),就能得到一条直线,其中
对t作图(图3-3a),就能得到一条直线,其中 ,是截距,
,是截距,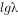 是斜率。
是斜率。
λ是种群离散增长模型中的重要参数,如果入>1种群上升,λ=1种群稳定,0<λ<1种群下降,入=0雌体没有繁殖,种群在下一代中灭亡。
②种群连续增长模型 在世代重叠的情况下,种群以连续的方式变化。这种系统的动态研究,涉及微分方程。 把种群变化率dN/dt与任何时间的种群大小联系起来,最简单的情况是有一恒定的增长率(per capita growth rate)r,它与密度无关,即
dN/dt=rN
其积分式为
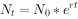
式中:e为自然对数的底。
例如,初始种群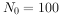 ,r为0.5/a,则一年后的种群数量为
,r为0.5/a,则一年后的种群数量为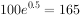 ,
,
二年后为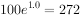 ,3年后为
,3年后为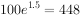 。
。
以种群大小对时间t作图(图3-3b),说明种群增长曲线呈“J”型。 但如
以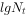 对t作图,则变为直线。
对t作图,则变为直线。

r是一种瞬时增长率(instantaneous rate of increase),很像复利的增长过程,如果r>0种群上升;r=0种群稳定;r<0种群下降。
种群一旦被证实为指数增长,则模型就有很大应用价值。例如,根据模型求人口增长率。1949年我国人口为5.41,1978年为9亿,求29a来的人口增长率。
解:
∵ 
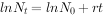
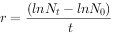
则:r=(1n9.5-In5.4)/(1978-1949)=0.0195,表示我国人口自然增长率为19.5‰,即平均每1000人每年增加19.5人。再求周限增长率λ。
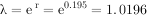
即每年是前一年的1.0196倍。
(2)与密度有关的种群增长模型:与密度有关的种群增长同样分为离散的和连续的两类。 与密度有关的种群连续增长模型,比与密度无关的模型增加了两点假设:
①有一个环境容纳量(通常以K表示),当 时,种群为零增长,即dN/dt=0;
时,种群为零增长,即dN/dt=0;
②增长率随密度上升而按比例降低。 最简单的是每增加一个个体,就产生1/K的抑制影响。
例如K=100,每增加一个个体,产生0.01影响,或者说,每一个体利用了1/K的“空间”,N个体利用了N/K“空间”,而可供种群继续增长的“剩余空间”只有(1-N/K)。这样,种群增长将不再是“J”型,而是“S”型。
“S”型曲线有两特点:①曲线渐近于K值,即平衡密度;②曲线上升是平滑的。
"S"型曲线的最简单数学模型是:

此即生态学发展史中著名的逻辑斯谛方程(logistic equation,或译阻滞方程)。
其积分式为:
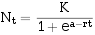
式中:a——截距,反映曲线对原点的相对位置,其值取决于N。。
逻辑斯谛曲线常划分为5个时期:①开始期,也可称为潜伏期,由于种群个体数很少,密度增长缓慢(a-b);②加速期,随个体数增加,密度增长逐渐加快(b-c);③转折期,当个体数达到饱和密度一半(即K/2时),密度增长最快(c-d);④减速期,个体数超过K/2以后,密度增长逐渐变慢(d-e);⑤饱和期,种群个体数达到K值而饱和(图3-4)。
逻辑斯谛模型的两个参数r和K,均具有重要的生物学意义。 如前所述,r表示物种的潜在增殖能力,而K则表示环境容纳量,即物种在特定环境中的平衡密度。虽然模型中的K值是一最大值,但作为生物学含义,它应该并可以随环境(资源量)改变而改变。

图3-4 逻辑斯谛曲线
逻辑斯谛增长模型的重要意义是:①它是许多两个相互作用的种群增长模型的基础;②它也是渔捞、林业、农业等实践领域中,确定最大持续产量(maximum sustained yield)的主要模型;③模型中两个参数r、K,已成为生物进化对策理论中的重要概念。
植物是定居的自养型生物,生长可塑性很大,这些特征直接影响了植物种群动态模型的建立,与动物种群动态模型的发展相比较,尚处于发展早期。
4.自然种群的数量变动
一种生物进入和占领新栖息地,首先经过种群增长和建立种群,以后可出现不规则的或规则的(即周期性的)波动,或较长期相对稳定;或有时出现骤然的数量猛增,即大发生,随后又是大崩溃;或有时出现长时期的下降,称为衰落,甚至灭亡。
(1)种群增长:自然种群数量的变动,从“J”型和“S”型增长均可以见到,但曲线不像数学模型所预测的光滑、典型,常常还表现为两类增长型之间的中间过渡型。例如,澳大利亚昆虫学家 Andrewartha 曾对果园中蓟马(Thrips imaginis)种群进行过长达14a的研究(图3-5)。 他发现,在环境条件较好的年份,其数量增加迅速,直到繁殖结束时增加突然停止,表现出“J”型增长;但在环境条件不好的年份则呈“S”型增长。 对比各年增长曲线,可以见到许多中间过渡型。

(2)季节消长:对自然种群的数量变动,首先要区别年内(季节消长)和年际变动。一年生草本植物北点地梅种群个体数有明显的季节消长(图3-6)。

8a间,籽苗数为500-1000株/m,每年死亡30%-70%,但至少有50株以上存活到开花结实,为次年种群的延续产出种子。 因此,各年间的成株数变动也相当少。
温带湖泊的浮游植物(主要是硅藻),往往每年有春秋两次密度高峰,称为“水华”(bloom)。其原因是:冬季的低温和光照减少,降低了水体的光合强度,营养物质随之逐渐积累;到春季水温升高、光照适宜,加之有充分营养物质,使具巨大增殖能力的硅藻迅速增长,形成春季数量高峰。 但是不久后营养物质耗尽,水温也可能过高,硅藻数量下降;以后营养物质又有积累,形成秋季较低的数量高峰。 这种典型的季节消长,也会受到气候异常和人为的污染而有所改变。掌握其消长规律,是水体富营养化预测和防治所必需的。

(3)不规则波动:我国是世界上具有最长气象记录的国家,马世骏(1965)从统计上探讨过大约1000a的有关东亚飞蝗(Locusta migratoria manilensis)危害和气象资料的关系,明确了东亚飞蝗在我国的大发生没有周期性现象(过去曾认为该种是有周期性的),同时还指出干旱是大发生的原因。1913-1961年东亚飞蝗动态曲线(图3-7)是各年发生级数变化序列图。通过分析还明确了黄、淮等大河三角洲的湿生草地,若遇到连年干旱,使土壤中蝗卵的存活率提高,是造成其大发生的原因。 但旱涝灾害与飞蝗大发生关系还因地而异。 据此他将我国蝗区分为4类,并分区提出预测大发生指标。 在对东亚飞蝗生态学的深入研究基础上,我国飞蝗防治工作取得了重大成就,已基本上控制了危害。

图3-7 1913-1961年东亚飞蝗洪泽湖蝗区的种群动态曲线(马世骏,1965)
(4) 周期性波动:经典的例子为旅鼠、北极狐的3~4a周期和美洲兔、加拿大猞猁的9~10a周期。 根据近30a资料,我国黑龙江伊春林区的小型鼠类种群,也具有明显3~4a周期,主要优势种为棕背平(Clethrionomys rufocanus),每遇高峰年的冬季就造成林木危害,尤其是幼林,对森林更新危害很大;并且,这3a周期与红松结实的周期性丰收相一致。 根据以鼠为主要食物的黄鼬(Nustela sibirica)的每年毛皮收购记录,证明黄鼬也有3a周期性,但高峰比鼠晚一年。有趣的是苏联远东阿穆尔地区森林脑炎发病率也有3a的周期性。 其原因是:鼠是革蜱的主要宿主,而革蜱是森林脑炎病毒的主要传播媒介。 紧随鼠高峰之后,次年出现革蜱的数量高峰和革蜱种群中脑炎病毒感染高峰,从而使林区中居民受革蜱叮咬而感染森林脑炎的患者增加。 因此,对鼠类种群数量动态的研究就成为森林脑炎流行病预测的重要指标之一。
(5)种群暴发或大发生:具不规则或周期性波动种群数量的生物都可能出现种群的大发生。最闻名的大发生见于害虫和害鼠。 例如蝗灾,我国古籍和西方圣经都有记载,如“蝗飞蔽天、人马不能行、所落沟堑尽平”等。非洲蝗灾至今仍有出现,索马里1957年一次蝗灾估计有蝗虫约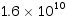 只之多,总重量达50 000 t。1967年我国新疆北部农区小家鼠大发生,估计造成粮食损失达
只之多,总重量达50 000 t。1967年我国新疆北部农区小家鼠大发生,估计造成粮食损失达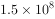 kg。
kg。
所谓赤潮是指水域中一些浮游生物(如腰鞭毛虫、裸甲藻、梭角藻、夜光藻等)暴发性增殖引起水色异常的现象,主要发生在近海,又称为红潮。 其主要成因是有机污染,氮、磷等营养物过多形成富营养化。其危害主要有:①藻类死体被微生物分解,消耗尽水中的溶解氧,鱼、贝等窒息而死;②有些赤潮生物产生毒素,杀害鱼、贝,甚至离海岸64 km的人,有时会受到由风带来毒素的危害,造成呼吸和皮肤的不适。
水生植物暴发造成危害的例子也不鲜见。一种槐叶萍(Salvinia molesta)原产巴西,1952年首次在澳大利亚出现,由于它每2.5d就能加倍,迅速增殖并扩散来,造成对交通、灌溉和渔业的严重危害。
(6)种群平衡:种群较长期地维持在几乎同一水平上,称为种群平衡。大型有蹄类、食肉类、蝙蝠,多数一年只产一仔,寿命长,种群数量一般是很稳定的在昆虫中,如一些蜻蜓成虫和具良好种内调节机制的昆虫(如红蚁、黄墩蚁),其数量也是十分稳定的。
在种群生态学发展史中,有的学者强调种群的变化或变动性,另一些强调平衡和稳定性。强调变动性的说,种群中始终有出生和死亡,在永恒不断变化之中,即使具有周期性的种群也不能具有绝对严格的周期。 强调稳定性的则说,各个种群一般均有一平均密度和平衡水平,当种群数量偏离此水平时,种群就有重新返回原有水平的倾向,即使具有剧烈数量波动特点的种类,如旅鼠和北极狐等,其种群的周期性(即几乎定期的升降)也可看做是一种稳定性。
(7)种群的衰落和灭亡:当种群长久处于不利条件下,或在人类过度捕猎、栖息地被破坏的情况下,其数量会出现持久性下降,即种群衰落,甚至灭亡。个体大、出生率低、生长慢、成熟晚的生物,最易出现这种情况。 例如由于第二次世界大战捕鲸业停顿,战后捕鲸船吨位上升,鲸捕获量上升,其结果导致蓝鲺鲸(Balaenoptera musculus)种群衰落,并濒临灭绝;继而长须鲸(B.physalus)日渐减少;就连小型的具有相当“智力”的白鲸(Delphinapterus)、海豚(Delphinus)、和鼠海豚(Phocaena)等也难逃厄运。
种群衰落和灭亡的速度在近代大大加快了,究其原因,不仅是人类的过度捕杀,更严重的是破坏野生生物的栖息地,剥夺了物种生存的条件。 物种的持续生存,不仅需要有保护良好的栖息环境,而且数量要达到最低种群密度。过低的数量因近亲繁殖而使种群的生育力和生活力衰退。
四、种群的空间格局
组成种群的个体在其生活空间中的位置状态或布局,称为种群空间格局(spatial pattern)或内分布型(internal distribution pattern)。种群的内分布型大致可分为3类:①均匀型(uniform);②随机型(random);③成群型(clumped)。(图3-8)。

均匀分布产生的主要原因是种群内个体间的竞争。例如森林中植物为竞争阳光和土壤中营养物,沙漠中植物为党争水分等。分泌有毒物质于土壤中以阻止同种植物籽苗的生长等。
随机分布中每一个体在种群领域中各个点上出现的机会是相等的,并且某一个体的存在不影响其他个体的分布。 随机分布比较少见,因为只有在环境资源分布均匀一致的情况下,或种群内个体间没有彼此吸引或排斥时才易产生随机分布。例如,森林地被层中的一些蜘蛛,面粉中的黄粉虫。
成群分布是最常见的内分布型。 成群分布形成的原因是:①环境资源分布不均匀,富饶与贫乏相嵌;②植物传播种子的方式使其以母株为扩散中心;③动物的社会行为使其结合成群。
内分布型的检验方法已有不少,如Greig-Smith(1952,1983)提出的等级方差分析法(hierarchical analysis of variance),Hill(1973)的3项轨迹方差法(three-term local variance analysis),Ripley(1978)的谱分析法(spectral analysis),杨持等(1984)的二维网函数插值法(two dimentional net function interpolation method)。最常用而简便的检验指标是方差与平均数的比率,即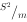 。其中,
。其中,
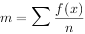
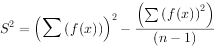
式中:x——样方中某种个体数;f ——含x个体样方的出现频率;n——样本总数。
如果把图3-8的均匀分布分成许多小方格,那么每方格中点数应该是相等的。对此进行取样和统计分析,因为各方格个体数相等,标准差等于零,所以=0。假如分布是随机的,则含有0,1,2,···的个体数的样方,其出现概率将符合泊松分布序列,所以=1。如果分布是成群的,含很少个体数(包括0个体在内)的样本,和含较多个体数的样本出现概率将比泊松分布的期望值高,因此,必然明显地大于1。
成群分布又可进一步按群本身的分布状况划分为均匀群、随机群和成群群,后者具有两级的成群分布。 由于种群空间分布格局反映着同一种群个体间的空间位置,无论是均匀群,还是随机群和成群群,既然有群,群与群之间就会有间隙。
为了刻画集群分布的特征,在研究中建立了格局规模、格局强度和格局纹理等属性概念。
格局规模:丛块大小以及丛块占调查面积的比例,叫做格局规模。
格局强度:丛块间的相对密度差或丛块内与间隙的相对密度差,称为格局强度。
格局纹理:丛块间空隙的大小则反映了格局纹理的粗细,丛块大且相隔远称为纹理粗糙,反之则称为纹理细密。
研究工作中,样地放置的位置和样地的大小都会影响到观察结果。其原因在于,如果某个种群呈随机群状分布,且格局纹理很粗,由于人的眼睛分辨不出来是什么格局,虽然放置样方是随机的,但仍可能把样方放置在群与群之间的间隙部位,或某一个群的内部,两种情况测定的结果不会是一样的,如果样方的面积足够大,有可能在这个大样方里包含了几个群和间隙,这个时候又会出现另一种结果。
种群空间格局研究是静态研究,比较适用于植物、定居或不大活动的动物,也适用于测量鼠穴、鸟巢等栖居所的空间分布。
构件生物的构件包括地面的枝条系统和地下的根系统,其空间排列是一重要生态特征,对个体的适应和生存具有重要意义。 例如在树冠层中,植物个体的枝叶系统的排列决定着光的摄取效率。 同样土壤中根分支的空间分布决定水和营养物的获得。 虽然枝条系统是“搜索”光的,根系统是“逃避”干旱的,但与动物依仗活动和行为进行搜索和逃避不同,植物靠的是控制构件生长的方向来寻觅养分并适应复杂多变的环境。


