第二节 种间关系
一、种间竞争
种间竞争(competition)是指具有相似要求的物种,为了争夺空间和资源而产生的一种直接或间接抑制对方的现象。在种间竞争中,常常是一方取得优势,而另一方受抑制甚至被消灭。
种间竞争的能力取决于种的生态习性、生活型和生态幅度等。具有相似生态习性的植物种群,在资源的需求和获取资源的手段上竞争都十分激烈,尤其是密度大的种群更是如此。植物的生长速率、个体大小、抗逆性及营养器官的数目等都会影响到竞争的能力。
(一)高斯假说
Gause(1934)首先用实验方法观察两个物种之间的竟争现象,他选择两种在分类上和生态习性上很接近的双小核草履虫(Paramecium aurelia)和大草履虫(P.caudatum)进行试验。对两个种取相等数目的个体,用一种杆菌为饲料,放在基本上恒定的环境里培养。开始时两个种都有增长,随后 P. aurelia 的个体数增加,而 P.caudatum 的个体下降,16 后只有P. aurelia 生存,而P.caudatum趋于最终的灭亡(图5-3)。这两种草履虫之间没有分泌有害物质,主要就是其中的一种增长得快,而另一种增长得慢,因竞争食物的结果,增长快的种排挤了增长慢的种。这就是当两个物种利用同一种资源和空间时产生的种间竞争现象。两个物种越相似,它们的生态位重叠就越多,竞争就越激烈。这种种间竞争情形后来被英国生态学家称之为高斯假说。
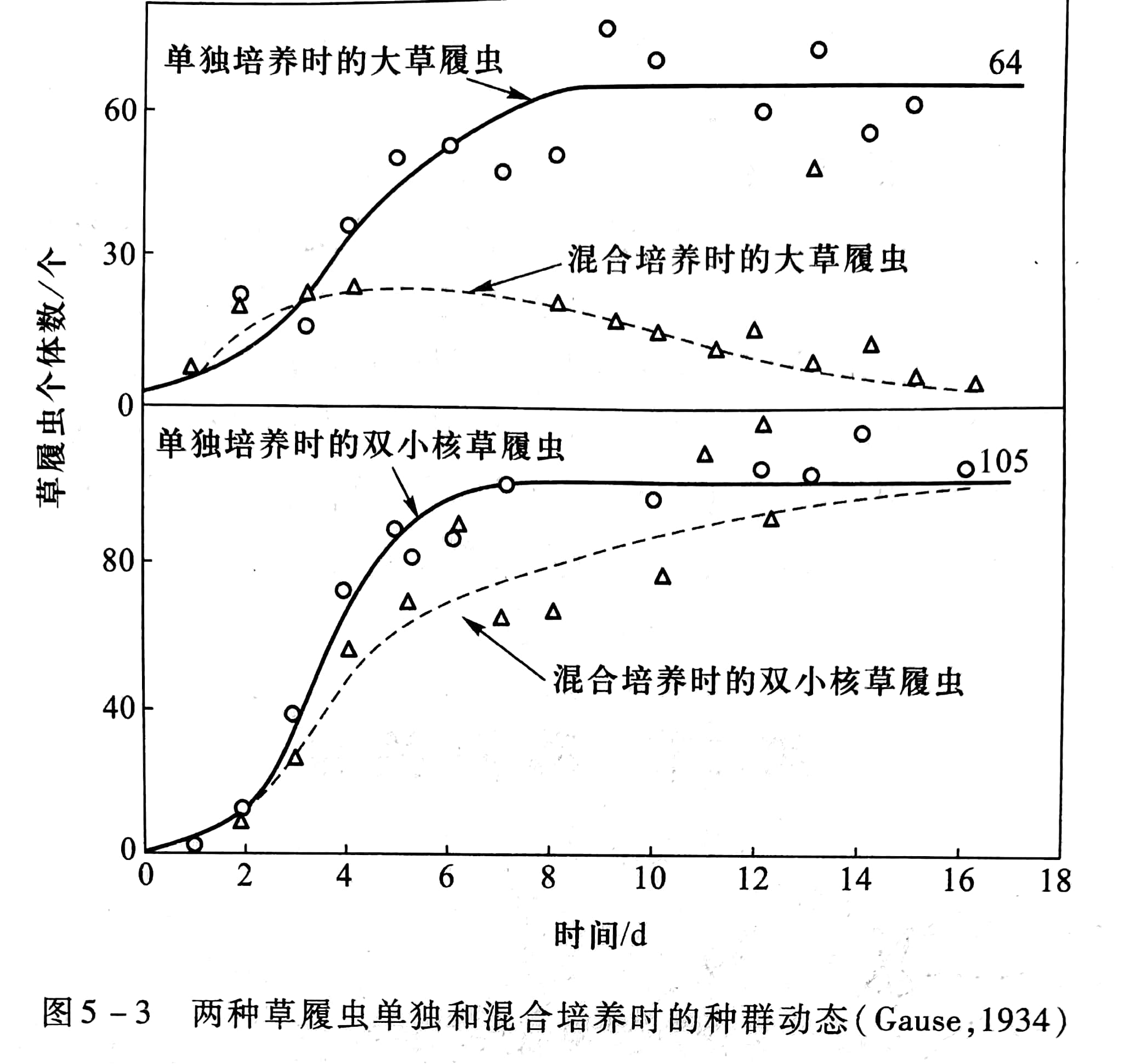
可以用竞争排斥原理(competitive exclusion principle)来表示这种概念,即在一个稳定的环境内,两个以上受资源限制的、但具有相同资源利用方式的种,不能长期共存在一起,也即完全的竞争者不能共存。其后,Park (1942, 1954)用赤拟谷盗(Tribolium castoneum)和杂拟谷盗(T.confusum)混养所做的实验,Tilman等(1981)用两种淡水硅藻和针杆藻(Synedraulna)所做的实验都得出了同样的结果。
(二) Lotka-Volterra模型
美国的Lotka(1925)和意大利的Volterra(1926)分别独立地提出了描述种间竞争的模型,是逻辑斯谛模型的引申。
现假定有两个物种,当它们单独生长时其增长形式符合逻辑斯谛模型,其增长方程是:
物种 1: 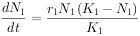
物种 2: 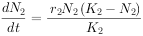
如果将这两个物种放置在一起,则它们就要发生竞争,从而影响种群的增长。设物种1和2的竞争系数为α和β(α表示在物种1的环境中,每存在一个物种2的个体,对于物种1种群的效应。β表示在物种2的环境中,每存在一个物种1的个体,对于物种2 种群的效应),并假定两种竞争者之间的竞争系数保持稳定,则物种1在竞争中的种群增长方程为:
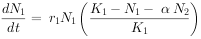
物种2在竞争中的种群增长方程为:

从理论上讲,两个种的竞争结果是由两个种的竞争系数 QB与K,、K,比值的关系决定的,可能有以下4种结果:
(1) a>K1/K2,和 β>K2/K1,两个种都可能获胜。
(2) a> K1/K2,和 β<K2/K1,物种1将被排斥,物种2取胜。
(3) a<K1/K2, 和 β>K2/K1,物种2将被排斥,物种1取胜。
(4) a<K1/K2, 和 β<K2/K1,两个种共存,达到某种平衡。
高等植物种群混合栽培或培养时所表现出的竟争结果都可以用 Lotka-Volt-erra 竞争方程来说明。
在温室中进行大麦和燕麦的竞争试验,在样方中以各种不同的比例进行播种,从纯大麦到纯燕麦种子,在生长季末收获种子,计算大麦/燕麦的输入比率和输出比率,结果见表5 -2。
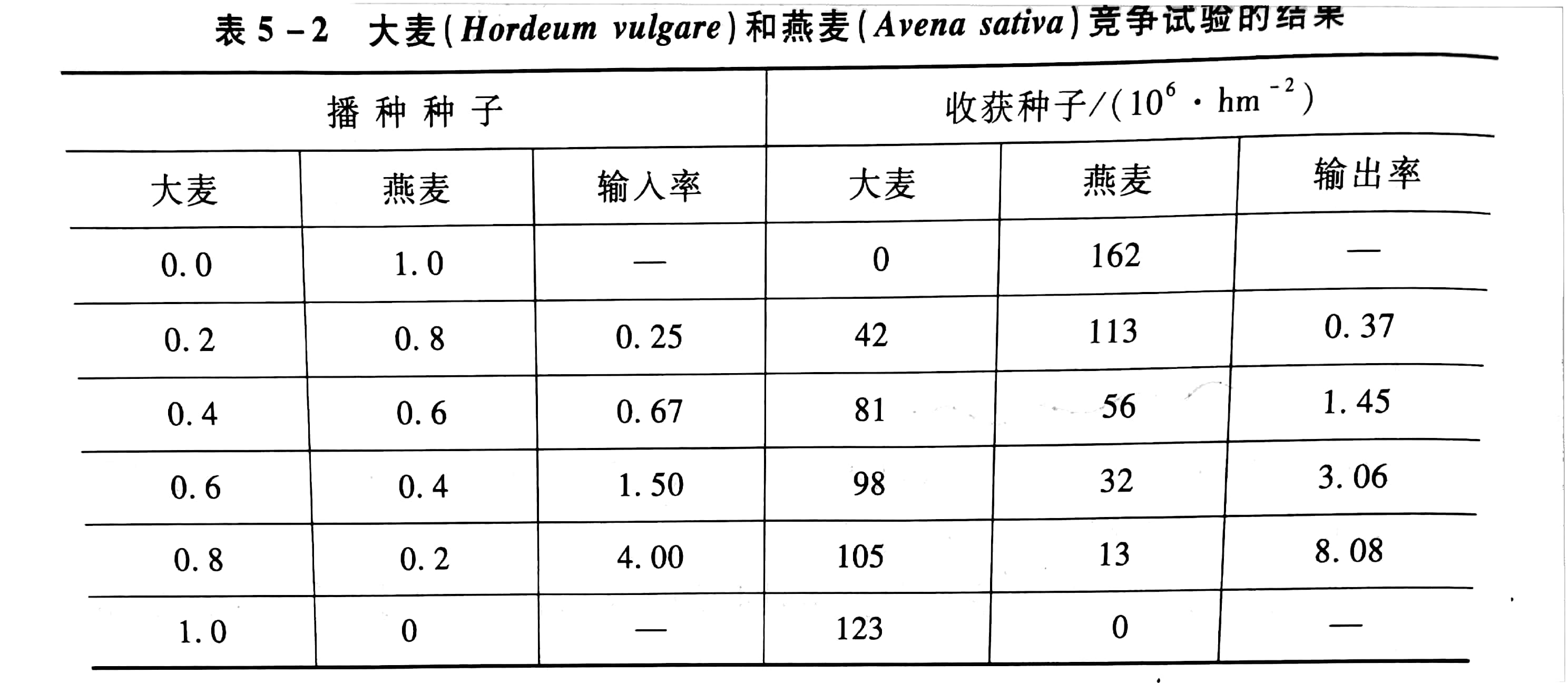
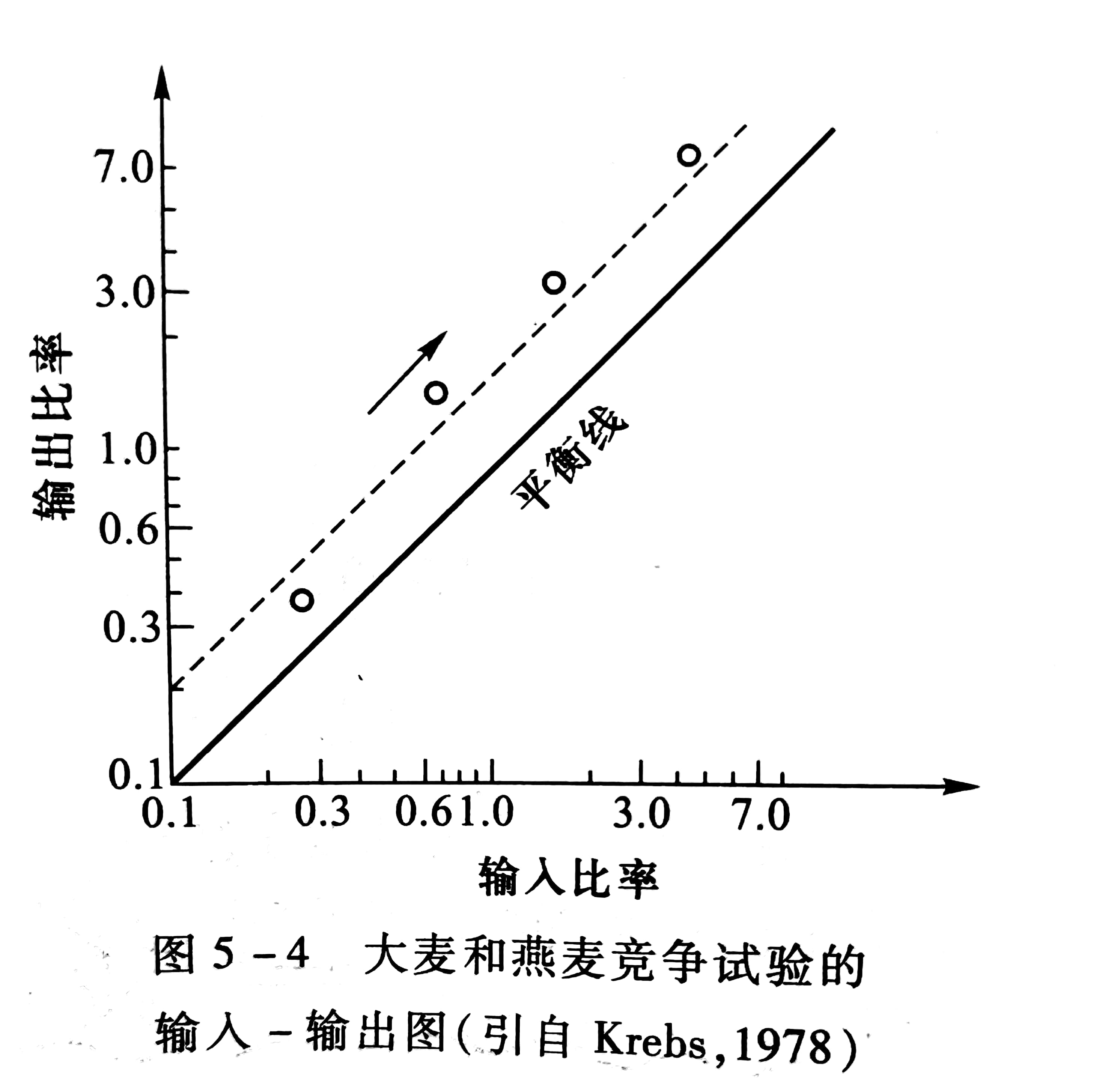
从表5-2中可见,虽然输入比率的大小不同,但是大麦的输出比率总是比输入比率大,因此,大麦是竞争中的胜利者。如果温室条件继续不变,大麦将最后把燕麦完全排挤掉,用输入、输出图解法分析,见图5-4。
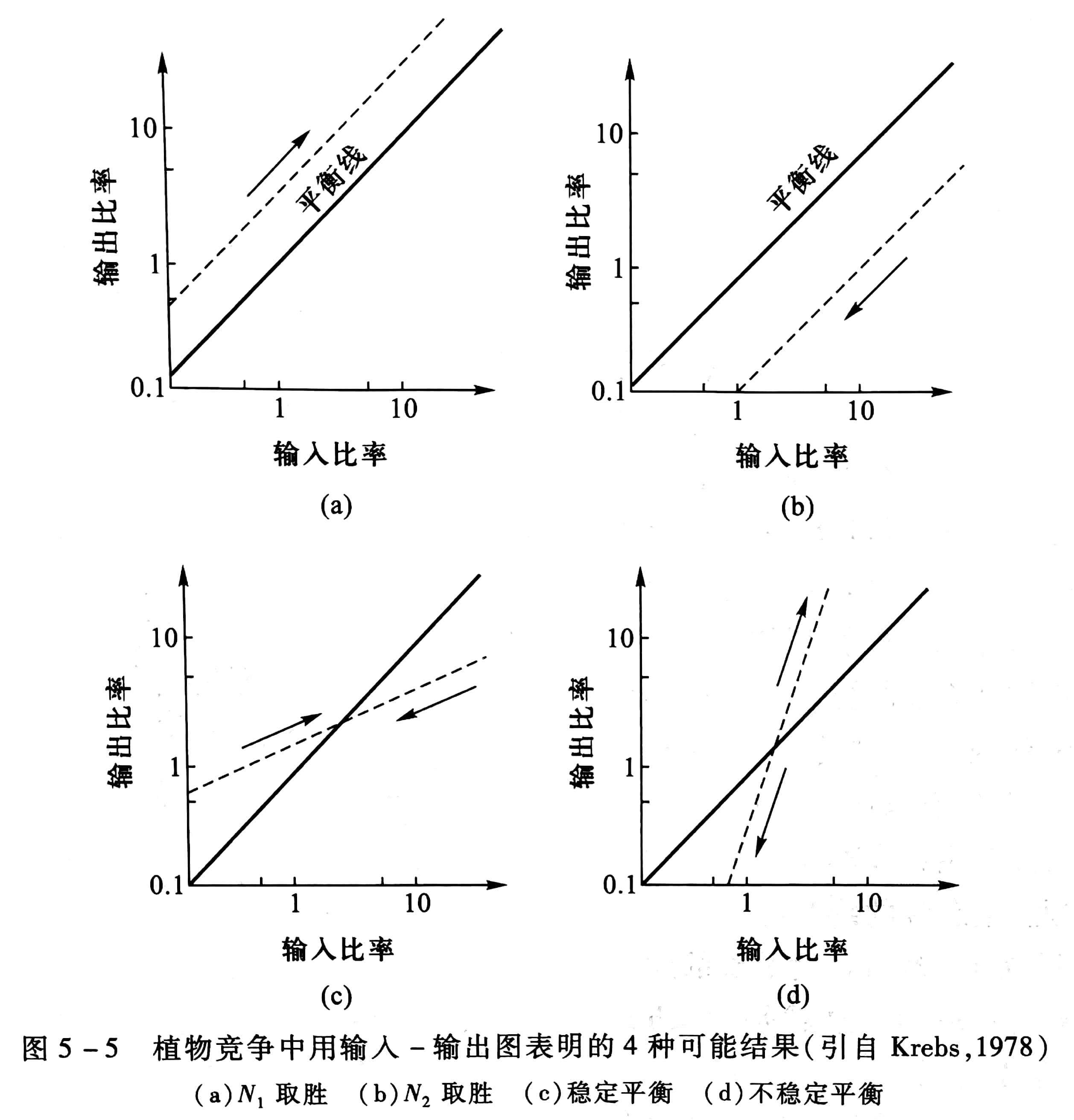
收获的种子可以再播种,从而成为下一年的输入比率,因此,试試验可连续进行若干代,然后将输出比率对输入比率作图(图5-5),可以有两种情况4 种结果:一种情况是某一种取胜,另一种被排挤。另一种情况是两个种可共存,或者是在某种稳定平衡下共存,或者是处于不稳定平衡中。
这样,通过一年生植物的竞争关系,可以提出一年生植物的竞争模型。模型根据播种时及收获时种子数的比例,即:
输入比率=物种甲播种的种子数/物种乙播种的种子数
输出比率=收货时物种甲种子数/收货时物种乙种子数
(三)生态位理论
生态位(niche)是生态学中的一个重要概念,主要指在自然生态系统中一个种群在时间、空间上的位置及其与相关种群之间的功能关系。明确这个概念对于正确认识物种在自然选择的进化过程中的作用、并在运用生态位理论指导人工群落建立中种群的配置等方面具有十分重要的意义。
生态位理论有一个形成与发展的过程。美国人J. Grinell (1917)最早在生态学中使用生态位的概念,用以表示划芬环境的空间单位和一个物种在环境中的地位。认为生态位“是一个种所占有的再细分了的环境”,实际上,他强调的是空间生态位(spatial niche)的概念。
英国生态学家C. Elton(1927) 赋予生态位以更进一步的含义,他把生态位看做是“物种在生物群落中的地位与功能作用”,强调的是物种与物种之间的营养关系。由于他把生态位概念的重点放在能量关系上,故实际上指的是营养生( trophic niche)。
英国生态学家 C. E. Hutchinson ( 1957) 发展了较现代的生态位概念,即n维生态位(n-dimensional niche)。他以种在多维空间中的适合性(fitness)去确定生态位边界,故对生态位的定义主要是指多维(超体积)生态位(multidimensional hypervolume niche),对如何确定一个物种所需要的生态位变得更清楚了。他还进一步提出基础生态位(fundamental niche)与实际生态位(realized niche)的概念,认为在生物群落中,能够为某一物种所栖息的理论上的最大空间为基础生态位,但实际上很少有一个物种能全部占据基础生态位,一个种实际占有的生态位空间为实际生态位。Hutchinson 的生态位概念目前已被广泛接受,现在让我们来做进一步的分析。
假如我们考察一个单一的环境因子(如温度)时,一个种将只有一个明确的适合度,亦即这个种只有在一定温度范围内才能生存和繁殖,这个范围就是这个种在一维上的生态位(图5-6a);假如我们同时考虑这个种在湿度上的适合范围时,生态位就成了两维的,并可以用面积表示(图5-6b);假如加上第三个环境因子(如食物颗粒大小),该生态位就成了三维的(图5-6c)。实际上,在生态系统中,物种的适合度受到许多生物和非生物因子的影响,因此,生态位的维数将大大多于3个,形成n- 维适合度明确的超体积(hypervolume)。这就是 Hutchinson 的生态位概念。

Hutchinson 的生态位概念中有如下一些重要的观点:
(1) 生态位是物种在群落中所处的地位、功能和环境关系的特性,它与生境(habitat)的含义不同,生境是指物种生活的环境类型的特性,如地理位置、海拔、水湿条件等。
(2) Hutchinson 将种间竞争作为生态位的特殊的环境参数,他把没有种间竞争的种的生态位称为基础生态位,这是物种潜在的可占领的空间,而把受竞争影响的现实的生态位称为实际生态位,其范围是由竞竞争因子所决定的。在这个概念中,Hutchinson 通过强调种间竞争对物种适合度的影响,表明基础生态位的一部分会由于竞争而失去。
(3) 生态位受生境的限制,生境会使生态位的部分内含失缺。
美国学者 R. H. Whittaker( 1970 )认为,生态位是一个种与群落中其他种相关联的位置关联的位置,即每个种在一定生境的群落中都有不同于其他种的自己的时间、空间位置,也包括在生物群落中的功能地位,他还指出,生态位的概念与生境和分布区的概念是不同的。分布区是指种分布的地理范围。生境是指生物生存的周围环境,生态位则说明在一个生物群落中某个种群的功能地位。
E. P. Odum(1971)将前人有关生态位的概念进行了综合,认为物种的生态位不仅决定于它们在哪里生活,而且亦决定于它们如何生活以及如何受到其他生物的约束。生态位概念不仅包括生物占有的物理空间,还包括它在群落中的功能作用以及它们在温度、湿度、土壤和其他生存条件的环境变化梯度中的位置。
不同的生物物种如动物、植物在生态系统中的营养与功能关系上各占据不同的地位,它们的生态位不同,由于环境条件的影响,生态位也会出现重叠与分化。不同生物在某一生态位维度上的分布,可以用资源利用曲线来表示,该曲线常呈正态曲线(图5-7),它表示物种具有的喜好位置及其散布在喜好位置周围的变异度。例如图5-7a 中各物种的生态位狭,相互重叠少,d >w,表示物种之间的种间竞争小;图5-7b 中各物种的生态位宽,相互重叠多,d<w,表示种间竞争大。
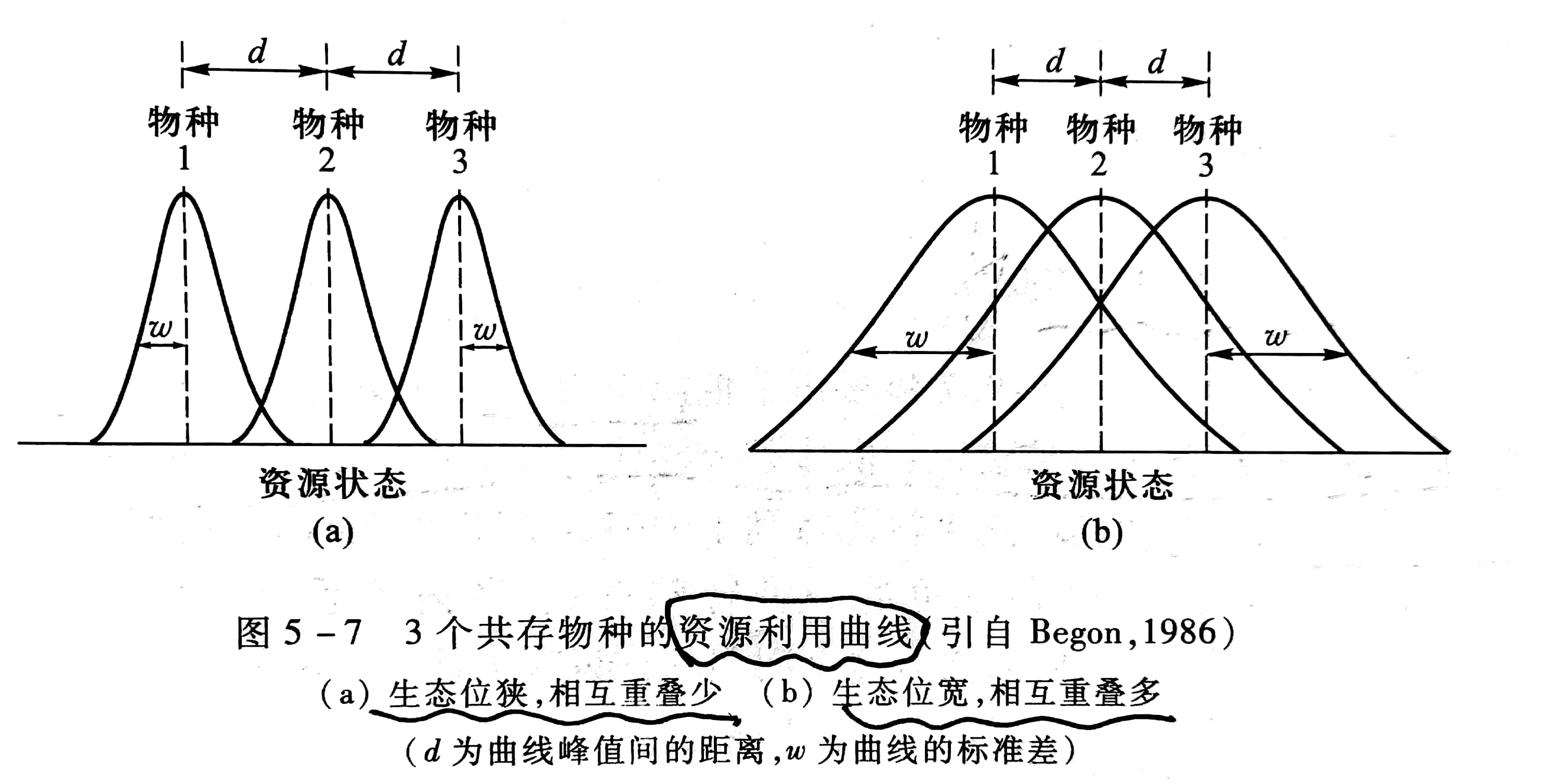
比较两个或多个物种的资源利用曲线,就能全面分析生态位的重叠和分离情形,探讨竞争与进化的关系。如果两个物种的资源利用曲线完全分开,则说明有某些未利用资源。扩充利用范围的物种将在进化中获得好处;同时,生态位狭的物种,如图5-7a,其激烈的种内竞争更将促使其扩展资源利用范围。由于这两个原因,进化将导致两物种的生态位靠近,重叠增加,种间竞争加剧。另一方面,生态位越接近,重叠越多,种间竞争也就越激烈;按竞争排斥原理,将导致某一物种灭亡,或者通过生态位分化而得以共存。后一种情形是导致两物种的生态位分离。总之,种内竞争促使两物种的生态位接近,种间竞争又促使两物种的生态位分离。
(1)具有不同分布区的种,其生态位往往是彼此分离的,彼此之间无竟争。(2)生活在同一地区的种,常常占据不同的群落生境,某些近缘种就这样被分隔开来,从而减少了竞争。除了地理分隔与群落分隔之外,两个物种的生态位还可因营养的选择吸收、个体大小、根系深沙、物候期等的不同,而彼此分隔开来,以减少竞争。
将前面讲述的竞争排斥原理与生态位概念应用到自然生物群落,则有以下一些要点:
(1)一个稳定的群落中占据了相同生态位的两个物种,其中一个种终究要灭亡。
(2)一个稳定的群落中,由于各种群在群落中具有各自的生态位,种群间能避免直接的竞争,从而又保证了群落的稳定。
(3)一个相互起作用的、生态位分化的种群系统,各种群在它们对群落的时间、空间和资源的利用方面,以及相互作用的可能类型方面,都趋向于互相补充而不是直接竞争。因此,由多个种群组成的生物群落,要比单一种群的群落更能有效地利用环境资源,维持长期较高的生产力、具有更大的稳定性。
二、他感作用
(一)他感作用的概念
德国学者 Molisch 于 1937 年提出了他感作用(allelopathy)的概念,他认为植物的他感作用就是一种植物通过向体外分泌代谢过程中的化学物质对其他植物产生直接或间接的影响。这个概念得到了大多数研究者的赞同。这种作用是种间关系的一部分,是生存竞争的一种特殊形式,种内关系也有此现象。在Molisch 的著作发表后,使人们对这个问题的认识有了长足的进展。
(二)他感作用的物质
20 世纪 40 年代以来,在植物他感作用的试验验证,克生物质的提取、分离和鉴定方面做了许多工作。Bode(1940)发现蒿叶的分泌物对毗邻植物具有明显的抑制作用,具有决定意义的成分主要是苦艾精——一种具有通式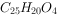 的化合物,多为一种芳香族的酸,他的研究后来曾为许多研究者所补充和证实。被鉴定出的叶分泌物还有:香桃木属(Myrtus)、桉树属(Eucalyptus)和臭椿属(Ailanthus)的叶面分泌物,主要是酚类物质, 如对羟基苯甲酸(hydroxy-cinnamic acid)、香草酸(vanillic acid)、阿魏酸(ferulic acid)等,它们对亚麻(Linum spp.)的生长具有明显的抑制作用。菊科植物 Encelia farniosa 是一种生长于美国加利福尼亚州南部半荒漠的多年生灌木,其叶分泌的一种苯甲醛物质对相邻的番茄、胡椒和玉米的生长有强烈的抑制作用,但对大麦、燕麦和向日葵的影响却很微弱。
的化合物,多为一种芳香族的酸,他的研究后来曾为许多研究者所补充和证实。被鉴定出的叶分泌物还有:香桃木属(Myrtus)、桉树属(Eucalyptus)和臭椿属(Ailanthus)的叶面分泌物,主要是酚类物质, 如对羟基苯甲酸(hydroxy-cinnamic acid)、香草酸(vanillic acid)、阿魏酸(ferulic acid)等,它们对亚麻(Linum spp.)的生长具有明显的抑制作用。菊科植物 Encelia farniosa 是一种生长于美国加利福尼亚州南部半荒漠的多年生灌木,其叶分泌的一种苯甲醛物质对相邻的番茄、胡椒和玉米的生长有强烈的抑制作用,但对大麦、燕麦和向日葵的影响却很微弱。
德国G. Grammer (1955)用 Allelopathy 的标题出版了专著,书中不但包括了高等植物之间的他感作用,而且包括高等植物与微生物之间、微生物和微生物之间、浮游动物之间等。他感作用的物质包括乙烯、香精油、酚及其衍生物、不饱和内脂、生物碱和配糖体等物质。
(三)他感作用的生态学意义
在自然界,植物一般均以群落的形式存在,从植物他感作用的角度来看,其种间结合的关系是形成群落的原因之一。
1.他感作用的歇地现象
植物他感作用的研究在农林业生产和管理上具有极重要的实践意义。在农业上,有些农作物不宜连作,连作则影响作物长势,降低产量。必须与其他作物轮作,连作时产生的这种现象,被称为歇地现象。例如早稻就是不宜连作的农作物,它的根系分泌的对-羟基肉桂酸,对早稻的幼苗起强烈的抑制作用,连作时则长势不好,产量降低。
红三叶草(Trifolium praterse)是繁殖力很强的牧草植物,它常形成较纯的群落,排挤其他的杂草植物。红三叶草含有多种异黄酮类物质,这些异黄酮类物质及其在土壤中被微生物分解而成的衍生物对其他植物的发芽起抑制作用,因而其他植物不易生长,同时也造成不宜连作的原因。
2.他感作用和植物群落中的种类组成
植物群落都由一定的植物种类组成,然而某种植物的出现会引起另一类植物的消退。Bode(1958)阐明了很久以来人们注意到的黑核桃 ( Juglans nigra)树下几乎没有草本植物的原因,因为该树种的树皮和果实含有氢化核桃酮(1,4,5 - 三羟基萘)。这种物质被雨水冲洗到土中,即被氧化成核桃酮,并抑制其他植物的生长。
银胶菊原产于墨西哥,是一种产橡胶的草本植物,它群生时,不但本身生长不好,而且对周围植物产生很大的影响,这是因为银胶菊植物根系分泌出反肉桂酸,抑制自身及其他植物生长。
3.他感作用与植物群落的演替
引起植物群落演替的原因很多,但大体上又分为外因和内因两大类,关于植物群落演替的内在因素,至今报道不多,目前认为他感作用是重要的因素之一。
北美加利福尼亚的草原,原来是由针茅(Stipa patahra)和早熟禾(Poa scabrella )等构成,由于放牧和烧荒,逐渐变成了由野燕麦和毛省麦(Bromusmollis)构成的一年生草本植物群落,以后又由于生长在这种群落周围的芳香性鼠尾草灌木(Sawia lencophylla, S. melifera)和蒿(Artemisia colifornica)的叶子分泌有樟脑和桉树脑等帖烯类物质,抑制了其他草本植物的生长,进而逐渐取代了一年生草本植物群落。在干旱季节,这些萜烯类物质聚集在土壤中,抑制了雨季时发芽的一年生植物。因此,在这些灌木的周围形成1~2m的草本植物不能生长的裸地。
近年来,植物他感作用的研究日益增多,研究方法更加先进,工作也更加深人,如利用气相色谱 - 质谱 - 计算机联用法,吸附浓缩技术以及火焰光度检查法等先进技术对克生物质的分离和鉴定。并倾向于研究他感作用的机制及在植物区系中的作用等方面。但相对而言,在苔藓,蕨类植物他感作用方面的研究较少。
三、捕食作用
(一)捕食者与猎物
生物种群之间除竞争食物和空间资源外,还有一种直接的对抗性关系,即一种生物吃掉另一种生物的捕食作用(predation ),生态学中常用捕食者(predator)与猎物或被食者(prey)的概念来描述,如食肉动物捕食食草动物或其他食肉动物,狮吃马,狼吃羊等。
由于捕食者与猎物的关系是在漫长的进化过程中形成的,因此捕食者可以作为自然选择的力量对猎物的质量起一定的调节作用。在自然界中,捕食者种群将猎物种群捕食殆尽的事例是很少的,被捕食的往往是体弱患病的或遗传特性较差的个体,淘汰了劣质,从而防止了疾病的传播及不利的遗传因素的延续。
在波兰,渔民们曾误认为水獭会把鱼类吃光,因此加紧捕猎水獭而使它们濒于灭绝。但很快又发现鱼类资源仍在不断减少,才知道水獭主要吃病鱼(因病鱼易被捕捉),因而也保持了种群良好的卫生状况。水獭被消灭却引起了鱼类传染病的蔓延和鱼类大量死亡。
捕食者具有捕食猎物的适应性。植食动物的牙齿常常具有很大的研磨表面,用以磨碎粗糙的植物组织。马的上下门齿有很强的对应性,使它们能咬断草的纤维茎。其他有蹄动物如牛、绵羊和鹿等虽然没有上门齿,但其下门齿能按一定角度把植物压到上颚处进行研磨。肉食动物的牙齿具有利于切割和撕咬的表面,既可将猎物把牢在口中又可把猎物撕裂成便于吞咽和消化的小块。许多捕食者都能够用前足抓住猎物并把它们撕成小块,如鹰、隼、猫头鹰和鸚鹉等就是田甘生有利爪的足和钩状喙制服和撕碎猎物。潜水鸟在捕食个体较大的鱼时,必须整个把鱼吞下去,因为其后肢专门用于游泳和潜水而不是用于抓握和拆解猎物。蛇虽然没有用于抓住猎物的附肢,但却生有可以扩张的颚能把很大的猎物整个吞下去。
猎物具有逃避捕食者的适应性。猎物逃避捕食的策略与捕食者的狩猎策略一样是多种多样的。隐藏、逃跑和积极自卫都属于防御,采取什么方法依捕食者——猎物关系的具体情况而定。对鹿、羚羊和其他食草动物来说,在广阔而平坦的草原上几乎找不到可以藏身的地方,因此它们只能靠及早发现捕食者和快速奔跑来求生。在能够隐藏或能在较安全的小生境中找到避难所的地方,动物常常对捕食者的存在极为敏感并能据此调整自己的行为。例如,在一些存在捕食性鱼类的池塘中,小鱼往往避开食物最丰富的开阔水域,大部分时间都生活在靠近水边的芦苇丛中。
在自然环境中,捕食者与猎物的关系是受许多因素影响的,往往是多种捕食者和多种猎物交叉着发生联系。如果捕食者是多食性的,它就可以选择多种不同的食物,给自身带来更多的生存机会,也具有阻止被食者种群进一步下降的重要作用。相反,就被食者而言,当它的密度上升较高时,可能会引来更多的捕食者,从而阻止其数量继续上升。例如在农田,猫头鹰多以鹌鹑为食, 当鹤鹑变少时,即转食啮齿类动物;在草原上,鼠类多的年份,食鼠动物如黄鼠狼、狐狸、鹰等也就有了充足的食物,可有效地阻止鼠类种群数目的持续上升。
(二)食草作用
食草作用(herbivory)是广义捕食的一种类型,其特点是被食者只有部分机体受损害,通常捕食者只采食植物的某一部分,留下的部分能够再生,例如牛羊吃草,植物本身没有逃脱食草动物的能力。当今的世界大部分是绿色的、植物没有被动物吃尽,其解释有二:① 食草动物在进化中发展了自我调节机制,防止作为其食物的植物都毁灭掉;② 植物在进化过程中发展了防卫机制。因此,食草动物与植物之间在进化过程中出现了选择竞争。
(1) 食物的质量能影响捕食者消化系统和排泄系统的适应性:植物含有长纤维分子如纤维素和木质素,它们形成了茎和叶的支持结构。与食肉动物的高蛋白食物相比,这些成分使植物更难于消化,因此很多食草动物(如兔和牛)的消化管都大大延长。很多食草动物的前肠部都有扩大的囊状区,而后肠就像发酵罐一样含有帮助消化的细菌和原生动物。由于具有这些适应性,食草动物才能使食物在消化管内停留更长时间并将其彻底消化。但这样的植食动物胃内必然会存留大量食物,这样就增加了其体重并降低了其活动性。
(2) 植物的补偿作用:植物在被动物啃食而受损害时并不是完全被动的,而是具有各种补偿机制。例如植物的一些枝叶在受损害后,自然落叶减少,而整株的光合率可能增强。受害植物可能利用储存于各组织和器官中的糖类得到补偿,或改变光合产物的分布来维持根/枝比的平衡。大豆如果在繁殖期受害,能以增加种子粒重来补偿豆英的损失。
(3) 植物的防卫反应:食草动物的“捕食”还可能引起植物的防卫反应,如产生更多的刺(机械防御)或化学分泌物(化学防御)。被牛啃食后的悬钩子的皮刺较未啃食过的长而尖;遭过钢蜂和树蜂危害的松树改变酚代谢,产生新的化学物;人工受伤的马铃薯和番茄能增加蛋白酶抑制物;荆豆顶枝在受到美洲兔的严重危害后,其枝条中会积累更多的毒素,变成兔所不可食的,这种化学保护可延续2~3年。
(4) 植物和食草动物的协同进化:协同进化(coevolution)是指在进化过程中,一个物种的性状作为对另一物种性状的反应而进化,而后一种的这一性状本身又作为前一物种性状的反应而进化的现象。在进化过程中,植物发展了防御机制,如有毒的次生物质,以对付食草动物的进攻。另一方面,食草动物亦在进化过程中产生了相应的适应性,如形成特殊的酶进行解毒,或者调整食草时间以避开植物的有毒化合物。
四、寄生与共生
(一)寄生
寄生(parasitism)是指一个种(寄生者)寄居于另一个种(寄主)的体内或体表,摄取寄主养分以维持生活的现象。寄生可分为体外寄生(寄生在寄主体表)与体内寄生(寄生在寄主体内)两类,在寄生性种子植物中还可分出全寄生与半寄生两类。全寄生植物从寄主那里摄取全部营养,而半寄生只是从寄主摄取无机盐类,它自身尚能行光合作用制造养分。全寄生的高等植物主要见于列当属(Orobanche)和菟丝子属(Cuscuta),半寄生植物有槲寄生(Viscum)和玄参科的植物等。
在植物之间的相互关系中,寄生是一个重要方面。寄生物以寄主的身体为定居的空间,并完全靠吸收寄主的营养而生活。因而寄生物使寄主植物的生长减弱,生物量和生产量降低,最后使寄主植物的养分耗竭,并使组织破坏而致死。因此,寄生物对寄主植物的生长有抑制作用,而寄主植物对寄生物则有加速生长的作用。
营寄生生活的高等植物,具有适应于寄生生活方式的形态解剖特征和生理特性。首先是寄生者的生物体简化,如槲寄生和小米草(Euphrasia)等半寄生植物它们仅保留含叶绿素的器官,能进行光合作用,但是水和无机盐类则从寄主植物体中获取。全寄生植物则含叶绿素的器官完全退化,如大花草(Raflesia arnoldii)、白粉藤属 (Cissus)是有花植物寄生者极端简化的例子,它们仅保留花,身体的所有其他器官都转变为丝状的细胞束,这种丝状体贯穿到寄主细胞的间隙中,吸取寄主植物的营养。
很多寄生植物还具有非常大的繁殖能力和很强的生命力,在没有碰到寄主时,能长期保持生活力,一旦碰到寄主植物,又能立即恢复生长,营寄生生活。如寄生在很多禾本科植物根上的玄参科独脚金属(Striga)植物,一株可产生 50 万个种子,可保持生命力 20 年不发芽,但一旦碰到寄主植物时,其种子就开始发芽生长,并侵人和寄生在寄主根中。
多数寄生植物只限于寄生在一定的植物科、属中,即寄生具有一定的专性,这类寄生植物为专性寄生植物,如菟丝子属和列当属中的很多种,常寄生在三叶草、亚麻、柳树、向日葵、大麻和宁麻等植物上。由于寄生具有一定的专性,故寄生者和寄主常常是协同进化的。
(二)偏利共生
共生(symbiosis)中仅对一方有利称为偏利共生(commensalism),附生植物与被附生植物是一种典型的偏利共生,如地衣、苔藓、某些蕨类以及很多高等的附生植物(如兰花)附生在树皮上,借助于被附生植物支撑自己,获取更多的光照和空间资源。藤壶附生在鲸鱼或螃蟹背上,并以其头顶上的吸盘固着在鲨鱼腹部等,都被认为是对一方有利、而对另一方无害的偏利共生类型。
(三)互利共生
互利共生(mutualism)是两物种相互有利的共居关系,彼此之间或者存在营养方面的相互依存,或者存在防卫性的相互照应。
营养方面的互利共生通常以双方在获取能量和营养时所特有的互补性为基础。地衣是藻类和真菌的共生体,藻类进行光合作用,菌丝吸收水分和无机盐,两者结合、相互补充,共同形成统一的整体生活在耐旱的环境中。菌根是真菌和高等植物根系的共生体。菌根有内生菌根和外生菌根两类,内生菌根是真菌菌丝穿人到高等植物的根部细胞里进行共生,很多高等植物如兰科、石楠科的帚石楠等都是内生菌根;外生菌根是真菌菌丝不伸人根部细胞里,而只是紧紧地包围在根外进行共生。外生菌根能增憎加根系的吸收面积,大多数乔、灌木树种如松树、云杉、橡树、山杨、榛树等都具有外生菌根。真菌从高等植物根中吸取糖类和其他有机物,或利用其根系分泌物,而又供给高等植物氮素和矿物质,两者互利共生。很多菌根植物在没有根菌时就不能正常生长或发芽,如松树在没有与它共生的真菌的土壤里,吸收养分很少,以致生长缓慢乃至死亡。在缺乏相应真菌的土壤上造林或种植菌根植物时,可以在土壤内接种真菌,或使种子事先感染真菌,便能获得显著的效果。同样,某些真菌如不与一定种类的高等植物根系共生,也将不能存活。
防卫上的互利共生表现为一方从另一方获得食物或隐蔽场所,同时回报给对方安全,如使其免受捕食和寄生物的攻击。在一些海洋生态系统中,专门有一些鱼和虾为其他鱼类清除体表和鳃上的寄生物,这些清洁夫以所清除的寄生物为食,而被清洁的鱼类则免除了寄生物的干扰。这种通常被称为清洁共生的关系在热带海域中最发达,在那里作为清洁夫的小鱼和虾往往具有鲜明的色彩,以吸引其他鱼类来接受其清洁服务。
动物与微生物之间互利共生的例子也有很多,如反刍动物与其胃内的微生物间形成了一种互利共生的关系,微生物既帮助了反刍动物消化食物,自身又得到了生存。白的肠道中生活着一种强厌氧性鞭毛虫(Tricho nympha),可消化纤维素。


