-
1 主页
-
2 参考
第三节 茎的初生生长及初生结构
一、顶端生长
在生长季,茎端分生组织不断分裂、伸长和分化,使茎的节数增加,节间伸长,同时产生新的叶原基和腋芽原基。这种由于茎端分生组织的活动而引起的生长,称为顶端生长(apical growth)。茎尖中进行的顶端生长的过程与结果依据细胞组织分区学说表解如下。
二、居间生长
禾本科、石竹科、石蒜科等植物保留在节间的居间分生组织进行的初生生长称居间生长(intercalary growth)。上述植物在进行顶端生长时,开始所形成的茎的节间基本不伸长,而是在节间留下一种初生分生组织(居间分生组织),植株发育到一定阶段,居间分生组织活动,形成初生结构。这种由居间分生组织进行的初生生长,称为居间生长。例如,小麦、高梁、玉米等禾本科植物的拔节。另外,在韭菜的叶基、花生的子房柄部位都存在有居间分生组织,可以引起韭菜割了一茬又长一茬、花生地上开花地下结实的现象。
三、双子叶植物茎的初生结构
茎端分生组织中的初生分生组织所衍生的细胞,经过进一步的细胞分裂、生长、分化而形成茎的初生结构,而初生结构不断积累的过程就是茎的初生生长。
茎为辐射对称的轴器官,茎的初生结构由表皮、皮层和维管柱三部分组成。
茎的三部分的详细结构如下(图3-10.图3-11 )。




(一)表皮
表皮位于茎的最外面,为原表皮发育而来的初生保护组织,通常由单层细胞组
成。有些植物茎的表皮细胞含花青素,因而茎呈红( 如红瑞木),紫等色。表皮细胞在横切面上呈长方形或方形,纵切面上呈长方形。蓖麻,甘蔗等的茎有时还有蜡质,可防止燕腾和增强表皮的坚韧性。旱生植物茎的表皮上,覆盖有角质层。
表皮中除表皮细胞外还常有气孔器,它是水分和气体出人的通道。此外,表皮上有时还有各种毛状体,包括分泌挥发油、黏液等的腺表皮。毛状体中较密的茸毛可以反射强光、降低蒸腾,坚硬的毛可以防止动物伤害,而具钩的毛可以使岂具攀缘作用。

dermis:真皮
dermal stem cell:真皮干细胞
melanocyte:黑素细胞

cutile:角质
hypodermis:下皮,真皮

(二)皮层
皮层位于表皮内方,是表皮和维管柱之间的部分,由基本分生组织分化而来。相比于根的初生结构的中皮层,茎的皮层要窄很多。
皮层包含多种营养组织。幼茎近表皮的薄壁组织细胞具叶绿体,能进行光合作用。水生植物的茎皮层细胞胞间隙发达,构成通气组织。有的植物茎皮层中还有分泌腔(棉、向日葵)、乳汁管( 甘薯)等分泌结构,有的含异细胞如晶细胞、单宁细胞(桃、花生)和石细胞群( 木本植物)。表皮内方一至数层皮层细胞常分化为厚角组织。在方形(薄荷、蚕豆)或多梭形(芹菜)的茎中,厚角组织常分布在四角或棱角部分( 图3-12)。



collenchyma:厚角组织
duct:导管

parenchyma:软组织
phloem:韧皮部
(三)维管柱
维管柱是皮层以内的中央柱状部分。双子叶植物茎的维管柱包括维管束。髓和髓射线等部分。
1.维管束
维管束是由原形成层分化而来,由初生木质部和初生韧皮部共同组成的束状结构。维管束在多数植物茎的节间成环状排成。双子叶植物的维管束在初生木质部和初生韧皮部之间有形成层,被称为束中形成层。束中形成层可以产生新的木质部和韧皮部,为无限维管束。
初生木质部由多种类型的细胞组成,包括导管、管胞、木薄壁组织和木纤维。导管和管胞属于输导组织,负责水和矿物质营养的运输。木薄璧组织由活细胞组成,具储藏作用。初生木质部的发育顺序为内始式: 原生木质部靠内,由环纹或螺纹导管组成,口径小; 后生木质部靠外,由梯纹,网纹或孔纹导管组成,口径大。
初生韧皮部由筛管、伴胞、韧皮薄壁组织和韧皮纤维共同组成,主要作用是运输有机养分。筛管属于输导组织,由筛管分子纵向连接而成。伴胞紧邻于筛管分子的侧面。柳皮薄壁细胞散生在整个初生团皮部中,常含有晶体、丹宁、淀粉等储藏物质。韧皮纤维常成束分布于初生韧皮部的最外侧。初生韧皮部的发育方式也为外始式(原生韧皮部靠外,后生韧皮部靠内)。
维管形成层位于初生韧皮部和初生木质部之间,在落的横切面上为排列整齐的扁平状组胞。
2.髓和髓射线
茎的初生结构中,由薄壁组织构成的中心部分称为髓(pith)。樟树等的茎髓部有石细胞。椴树等的茎髓外方有小型的厚璧细胞,国绕着其内侧的大型细胞,者界线分明,这个外围区称为环髓带。还有的植物髓中有异细胞( 如品细胞、单宁细胞、黏液细胞等)生于薄壁细胞之间。伞形科、葫芦科等植物茎的髓部成熟较早,随着茎的生长,节间部分的髓被拉破,形成的空腔称为髓腔。例如,胡桃、枫杨等的茎,在节间还可看到残存的片状髓。
Pith, or medulla, is a tissue in the stems of vascular plants. Pith is composed of soft, spongy parenchyma cells, which in some cases can store starch. In eudicotyledons, pith is located in the center of the stem. In monocotyledons, it extends also into flowering stems and roots. The pith is encircled by a ring of xylem; the xylem, in turn, is encircled by a ring of phloem.
Pith
The central cylinder of a twig –the pith — is composed of non-woody tissue having a markedly different texture than the surrounding remainder of the twig. Viewed in a longitudinal section made with a razor blade or sharp knife, there are several potentially distinctive aspects of pith. Most species have pith that is fairly unremarkable: smooth without any interuptions, called continuous homogenous pith. More interesting pith has cross walls (partitions). If these partitions occur additional to continuous material in the center of the twig the pith is said to be diaphragmed, whereas if the partitions occur in an otherwise hollow space the pith is chambered. A few species lack pith altogether.
 Pith examples (Left to Right): Continuous homogenous pith of tree-of-heaven (Ailanthus altissima).
Pith examples (Left to Right): Continuous homogenous pith of tree-of-heaven (Ailanthus altissima).
2. Diaphragmed pith of tulip-tree (Liriodendron tulipifera)鹅掌莲.
3. Chambered pith of black walnut (Juglans nigra).胡桃木
4. Hollow (no) pith of honeysuckle (Lonicera sp.).金银花
髓射线(pith ray)位于维管束间,为连接髓和皮层的薄壁组织,由基本分生组织发育而来,也称初生射线。在横切面上星放射状,起横向运输的作用。髓射线和髓均具有储藏营养物质的功能。
以上所述是茎节间部分的初生结构,而茎包括节间和节两部分。节的结构较复杂。节部着生叶,叶内的维管束通过节部和茎内维管束相连,叶片和腋芽分化出来的维管束都在节上转变汇合,具体过程将在茎和叶的联系中详述。


Stem of a dicotyledon plant with only primary tissues. The ground tissue system includes the pith and cortex and the vascular tissue system is in discrete bundles.
四、单子叶植物茎的初生结构
(一)单子叶植物茎节间的初生结构
单子叶植物和双子叶植物的茎在结构上有许多不同。大多数单子叶植物的茎,只有初生结构,少数的有次生结构。
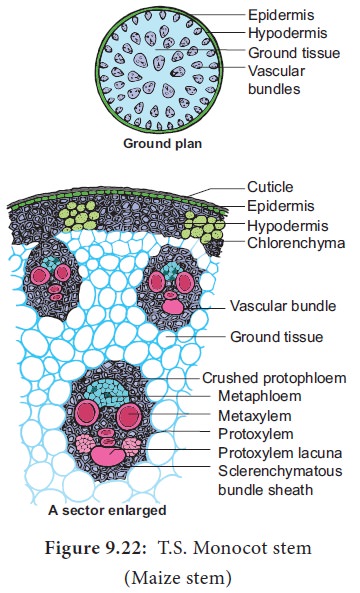
绝大多数单子叶植物茎的维管束仅由木质部和韧皮部组成,为有限外韧维管束。维管束有两种排列方式: 玉米(图3-13)、高粱、甘蔗等的维管束无规则地散生于基本组织内,外多内少,皮层和髓不易分辨; 水稻(图3-14)。小麦等的维管束一般为两圈,中央为髓(但在苦长大时,髓部破裂形成髓腔)。

以禾本科植物长米的器为代表,说明一般单子叶植物茎节间的初生结构特点。玉米成熟茎的节间部分,在横切面上可以明显地看到表皮、基本组织和维管束3 个部分。
1.表皮
表皮位于茎的最外方,细胞排列整齐。茎的表皮细胞有长短之分,长细胞夹杂着短细胞。长细胞较多且角质化。短细
胞位于长细胞之间,分为两种: 木栓化的栓质细胞和含有二氧化碓的硅质细胞。此外,表皮上还有少量气孔器( 图3-15)。

2.基本组织
除与表皮相接的部分外,整个基本组织均由薄壁细胞组成,越向中心,细胞越大,维管束散生其中。基本组织近表皮的部分由厚璧细胞组成,可增强茎的支持功能。幼嫩的茎,在近表面的基本组织细胞内,因含有叶绿体而量绿色,能进行光合作用。
3.维管束
玉米茎内的维管束散生在基本组织中,在横切面上星近卵圆形,外面是厚壁组织组成的鞘状结构,称为维管束鞘( 3-16)。


木质部和韧皮部内外排列,为有限维管束。
韧皮部中的后生韧皮部,细胞排列整齐,在横切面上可以看到多边形(六角形、八角形)的筛管细胞和交叉排列的长方形伴胞。在韧皮部外侧和维管束鞘交接处,可以看到由于后生韧皮部形成而被挤压的原生韧皮部。
木质部位于韧皮部之内。紧接后生韧皮部的部分,是后生木质部的2 个较大的孔纹导管。向内是原生木质部,由2-3 个口径较小的环纹导管或螺纹导管组成。维管束的2 个孔纹导管,和直列的环纹或螺纹导管,构成V字形结构,这是不本科植物茎中较明显的结构。原生木质部中直列的2 个或3 个导管,有时可能只存在1个或2 个,最里面的1个被腔隙替代,这是由于环纹或螺纹导管在生长过程中被拉破造成。从以上的结构中,可以看出,维管束中韧皮部的分化,是由外向内,即外始式。而木质部的分化,是由内向外,即内始式。
(二)单子叶植物茎节的结构
以禾本科植物为例,说明单子叶植物茎节的结构。禾本科植物的茎和叶鞘相连处形成了节部。在内部结构上,由于上端的节间维管束以及从叶鞘延伸进入的维管束(叶迹)在此交织汇合,出现了较复杂的结构。将小麦的茎,由上至下,从上部节间经节部,再到下部节间作连续切片观察,可以看到这种维管系统汇合排列的变化过程(图3-17)。


小麦茎的节间中空,在节部成为实心。叶鞘在较高水平上向一边开放,到了靠近节部成为封闭的。在内部结构上,茎节上面的维管束成横向和斜向的分布,而在节内及节的下面,就重新组合排列到较外面的周围。在叶鞘和茎连接处的下面,较小的叶迹延伸到茎轴的外围,较大的叶迹则变成了茎轴的中间维管束柱的部分。