★知识框架(必看)↓↓↓


★章节视频(必看)↓↓↓
★章节PPT↓↓↓
★章节文字稿↓↓↓
轮虫以单性生殖方式,繁殖群体数量迅速增长。人工培育轮虫的目的是产生大量的生物饵料。因此,维持单性生殖方式是最理想的。而两性生殖的出现,则意味着轮虫群体数量增长减慢,甚至在产生大量休眠卵后虫体大量死亡,造成培养的崩溃。但,为什么我们又说轮虫休眠卵的产生对生产是具有积极意义呢?
休眠卵产生对轮虫和生产的积极意义何在?又是什么触发了轮虫产生休眠卵呢?
首先,我们来解决第一个问题:休眠卵产生对轮虫和生产具有什么积极意义?
轮虫休眠卵从母体排出沉入水底或留在死亡后的母体内,大小和夏卵相近,卵壳很厚,不透明,形状多样,壳上常长有壳纹和长短不一的刺。
休眠卵的这些特点,使其在休眠期间,可以抵御外界高度的干燥、低温和高温,以及其他化学因子的剧烈变化等,恶劣的环境条件。而当温度、食物、PH、容量等外界环境条件适合时,休眠卵解除滞育状态,又可以孵化形成子代新个体,进入单行生殖世代。所以说,轮虫有性生殖产生休眠卵,以渡过不良环境,不仅对维持其种族的生存、繁殖有极重要的生物学意义,也是人工轮虫保种的重要途径。正是因为轮虫休眠卵的抗逆性强,与轮虫相比,其更便于运输。
休眠卵的产生需经过5个步骤,随着轮虫孤雌生殖的进行,由非混交雌体产生混交雌体;未受精的混交雌体产生雄轮虫;雄轮虫与混交雌体交配;受精;受精后的混交雌体产生休眠卵。是什么原因使得孤雌生殖非混交雌体产生混交雌体呢?
目前已知,一系列内源性和外源性因素皆可通过影响轮虫种群中混交雌体的百分率和受精率,以及受精的混交雌体的产卵量,从而影响休眠卵的形成。
在要了解休眠卵的产生,要从以下4个方面进行了解,分别是:诱发两性生殖的因子,混交雌体的受精率,受精的混交雌体的产卵量,影响休眠卵形成效率的因素。
我们首先来了解,诱发两性生殖的因子。混交雌体的产生是轮虫,由孤雌生殖向两性生殖转变及休眠卵形成的第1步,它受众多的外源性及内源性因素的影响和制约。
已确定的外源性因素主要有:生育酚;种群密度;食物;温度;光照;盐度;pH。内源性因素主要有:遗传因素;母体的年龄;轮虫孤雌生殖的累积世代数。
α-生育酚可直接作用于轮虫非需精卵胚胎发育的后期,而诱导混交雌体的形成。。
密度的自我调节是种群为保持稳定的一个重要特点,生物为了其种群的自身生存与发展,在环境条件改变时,各种不同形式的调节方式就开始起作用了。对于具有世代交替现象的轮虫来讲。在适合的环境下,以快速、便捷的孤雌生殖扩大种群,迅速占据空间,对种群十分有益。当种群密度接近环境最大负荷时,种群开始出现雄性个体和混交雌体交配,进行有性生殖,这种方式无疑使得轮虫以休眠卵形式离开种群。使密度得到控制。
食物的种类、适口性、特有成分、生长状况、年龄、浓度对轮虫混交雌体的形成具有明显的诱导作用。
温度对轮虫混交雌体形成的百分率有显著影响,尤其是不可预测的骤然变化,对混交雌体形成具有明显的刺激效应。光照的昼夜光长变化、光谱成分和光照强度对轮虫混交雌体的形成具有诱导作用。盐度是影响咸水性轮虫混交雌体形成的主要外源因子之一。 pH的高低直接影响轮虫混交雌体的产生,较高的pH环境下培养轮虫时,其混交雌体产生的百分率相对较高。。
混交雌体的形成常因同种轮虫,不同品系间的遗传差异而存在较大的差异,遗传为决定性因素,环境因子对混交雌体更多的是调节作用。
同时,母体的年龄与轮虫混交雌体的形成有密切的关系。有研究发现,萼花臂尾轮虫和红臂尾轮虫母体的年龄,与其后代中混交雌体的比例呈显著的负相关。
最后,轮虫有性生殖至少经两阶段完成:第1个阶段为获得产生混交雌体的能力,这种能力往往在轮虫孤雌生殖进行一代后即可自然形成。第2个阶段是外源性因子,如高种群密度的作用,这种作用只有对经过了第一阶段的轮虫才有效。因此,轮虫孤雌生殖世代数的积累对轮虫混交雌体的形成具有诱导作用。
诱发轮虫两性生殖的因子,就介绍到这里。接下来我们学习混交雌体的受精率。
即使当种群混交雌体被诱导产生,并随之出现雄性个体,受精作用也并非一定会发生,休眠卵也并非一定会形成。就轮虫本身而言,受精作用的发生主要与以下几个方面有关:雌雄相遇及交配的可能性;雄轮虫的受精力;雌体是否易于受精。
我们先来看第一个方面,雌雄相遇及交配的可能性。雌雄相遇的可能性与种群密度有关。但,交配的可能性则取决于,雄体对雌体产出的一组与交配有关的糖蛋白的感受能力。
同时,雄轮虫的受精力也会影响混交雌体的受精率。雄轮虫的年龄与其受精力有关,随着雄轮虫年龄的增大,精子的数量和运动能力降低,从而导致雄轮虫的受精力降低。母体的摄食对子一代雄体的受精力也起着重要的作用,混交雌体在轮虫种群的对数增长期所产的雄体,比在稳定期产的,具有较多的精子数量和较强的运动能力。
最后,影响混交雌体受精率的还有雌体是否易于受精。雌体欲被受精,就必须吸引雄体并进行交配。雌雄相遇并进行交配的可能性的34%取决于雌体的吸引力。不同品系间雌体对雄体的吸引力不同。
雌体是否易于受精也与雌体的年龄有关。如,西氏晶囊轮虫的混交雌体只在出生后的4小时内可与雄体发生受精作用,褶皱臂尾轮虫在出生后的9小时内可与雄体发生受精作用。
以上便是影响混交雌体受精率的主要因素。接下来我们进入轮虫休眠卵产生的第三个内容,受精的混交雌体的产卵量。
环境因子对轮虫受精的混交雌体的产卵量有明显的影响。温度、碱度和食物的种类是重要的环境因素。
温度升高使受精的混交雌体的产卵量增大。
在碱性条件下,萼花臂尾轮虫受精的混交雌体在pH为8.5时的平均产卵量最高。
以斜生栅藻、蛋白核小球藻、斜生栅藻与蛋白核小球藻混合三种方式投喂培萼花臂尾轮虫,发现,单投喂斜生栅藻、投喂斜生栅藻与蛋白核小球藻的混合料两种投喂方式均能显著地提高休眠卵的产量。
最后,我们在用一点时间来学习与轮虫休眠卵产生有关的最后一个内容——影响休眠卵形成效率的因素。目前已知影响休眠卵形成效率的因素主要有:遗传因素、温度、食物种类和浓度、培养液的盐度、pH等。如何提供有利的条件,提高休眠卵形成效率,则因种而异。在生产中需要进行调节时,应根据轮虫的种类和品系来定,这里我们不再一一展开。
★知识框架(必看)↓↓↓
★章节视频(必看)↓↓↓
★章节PPT↓↓↓
★章节文字稿↓↓↓
休眠卵的产生对人工轮虫培养有十分重要的意义,因为轮虫一般采用保存休眠卵的方式进行保种。
如何获得足量的休眠卵,如何估测休眠卵的数量,如何有效保存休眠卵,将是下次人工轮虫培养成功的前提。
下面我们将从休眠卵的诱发,休眠卵的采集、分离和定量,休眠卵的保存三个方面学习轮虫休眠卵的相关知识。
1、休眠卵的诱发:对诱发轮虫两性生殖的因子已在上一节中介绍。目前在生产中诱发两性生殖形成休眠卵的方法,是以高的种群密度和饥饿刺激相结合进行。即先以常规方法培养轮虫,不采收,待轮虫数量达高峰时,突然停止投喂饵料并停止充气,池塘中饵料很快耗尽,即会出现两性生殖产生大量休眠卵。其次,还可以改变盐度或水温,促进轮虫产生休眠卵。把低温条件下培养的轮虫放到加热至29~30℃的培养液中,每两天反复操作一次。通过温度刺激可以人为的促使轮虫产生大量休眠卵。
2、休眠卵的计数:轮虫休眠卵的采集可以使用有机玻璃采泥器,即休眠卵采集器,该采集器的形状为长圆筒形,体积为600立方厘米。
休眠卵的采集方法为:在每个池设2~4个采集点,将采集器垂直插入池塘底泥中,采取约10厘米厚的底泥,并切去采集器下部5厘米的底泥,留上部5厘米的底泥装于1000ml的盛泥钵中进行下一步的分离。
将底泥装于1000ml的盛泥钵后,加水稀释到600ml,搅拌均匀制成泥浆水。
抽搅拌均匀后的泥浆水10ml,注入50ml的烧瓶中等到分离。
分离前先在用精盐调成的饱和食盐水中,加入精盐质量20%的蔗糖,配制成糖盐高渗液。
分离第一步,将高渗液徐徐加入上述盛泥浆水的烧瓶中,用玻璃棒搅动1分钟,静置20分钟。
分离第二步,等泥沙下沉后,再次使用玻璃棒搅动,并再次加入高渗液,使液面略突出于瓶口,这时休眠卵逐渐上浮,静置20分钟后即可计数。
计数需要用的工具主要有浮游生物计数框、光学显微镜;
计数的操作如下:首先,用浮游生物计数框在突出瓶口的液面上黏取上浮休眠卵,一般连取3~4片即可将上浮休眠卵黏取尽;随即,将计数框置于光学显微镜下,用低倍镜观察并计数。
显微镜下统计的数量为样本液的休眠卵总数,因此我们还需要使用公式计算出整个池塘的轮虫休眠卵密度。池塘休眠卵密度等于被稀释后的泥水体积,乘于显微镜下观察到的休眠卵数量,除于所取的稀释后的泥浆水体积,再除于采泥器底面积。根据分离操作,我们可知被稀释后的泥水体积为600ml,所取的稀释后的泥浆水体积为10ml。所以公式也可以简化为:池塘休眠卵密度等于60乘于显微镜下观察到的休眠卵数量,除于采泥器底面积。
3、休眠卵的保存:轮虫是采用保存休眠卵的方式保种的,因此,保存休眠卵是轮虫培养中必不可少的工作。轮虫休眠卵保存的方法主要有三种:第1种是冰箱保存,第2种是干燥保存,第3种是原池保存。
冰箱保存休眠卵一般使用到的用品有:带软胶塞棕色玻璃瓶、冰箱。操作的流程为:收集分离休眠卵,将收集分离获得的休眠卵直接装入带软胶塞棕色玻璃瓶中,盖上盖子放于5℃冰箱内保存。可保存1~2年之久,甚至有人发现冰箱保存了8年以上的休眠卵仍保持有孵化能力,可见休眠卵耐受恶劣环境的时间相当长。
在轮虫冰箱保存休眠卵的操作中,较为复杂的便是休眠卵的收集分离工作。在生产中,休眠卵的收集分离主要有三种方式:方法1,从富含休眠卵的轮虫冷冻品中直接分离筛选;该来源的休眠卵分离的方法为,将富含休眠卵的轮虫冷冻品放在容器内自然融化,容器容量为轮虫冷冻品的5倍以上,等完全融化之后,将容器加满自来水稀释,用木棍用力搅动溶液,可以正反方向交替搅动共计10min,静止10min后轻轻撇去上层浮沫。之后再加满水稀释,按上述方法反复操作若干遍,直到静止后没有浮沫(或极少浮沫存在),再把容器中的液体轻轻倒人另外一个同样大小的容器,底层的沉积物废弃掉。为了保证收集休眠卵的纯度,可以每问隔10min再倒--次容器,每次倒容器之前撇沫,倒容器时将底层沉积物废弃,连续操作三次,轮虫休眠卵的纯度可达到90以上,最后用300目筛绢做的网袋,将容器内的液体浓缩收集,就是轮虫休眠卵。
方法2,秋冬季节到养殖轮虫的池塘或轮虫资源丰富的水坑中,排水刮取底泥,再进行分离筛选。该来源的休眠卵分离的方法为,将捞取的富含轮虫卵的底泥,放入一个容器内,加满自来水轻轻搅动,待基本均匀后静止10min后,用8O目筛绢做的捞网将上层悬浮物撇除,之后将悬浊液轻轻倒入另外一个同样大小的容器,底层的沉积物废弃。如此操作三遍左右,基本可以保证悬浊液中的轮虫休眠卵纯度达90以上,最后同方法1一样进行浓缩获取休眠卵。
方法3,捞取正在繁殖高峰期的轮虫,放人室内水泥池(也可以是其他容器)进行人工诱发休眠卵的产生,再进行分离纯化。人工诱发的操作一般为:当室内水泥池轮虫达到一定的高密度时进行饥饿或降温、降盐等理化环境变化刺激,很快会产生大量休眠卵,气温15~25℃时,一般2~3d即达到可收集水平,气温15℃以下可以适当延长时间(以检查底层休眠卵数量为准)。收集时,排水至250px,搅动底层水,在排水口安装300目筛绢做成的网袋以接收排出的休眠卵和杂质,然后开始排水,然后将收集的卵和杂质按方法2的分离步骤进行操作,即可得到休眠卵。
干燥保存的操作则是,将收集分离的轮虫休眠卵,倒在筛绢框(300目以上)或报纸上,直接晾干或用冷风吹干(不可太阳直射),干燥后的轮虫休眠卵呈薄片状,土黄色,装入棕色玻璃小瓶置于4℃冰箱中保存。但这种储存方法轮虫休眠卵的孵化率随着时间的延长而降低,干法储存1年,孵化率下降50%,但继续冰储存至两年,孵化率不再明显下降。
休眠卵也可以不吸出,保留在原培养容器或原池中,原池水不能排去,也不能更换新水。若需要再培养时,把原始水排掉大部分,注意保留底部休眠卵,换入新鲜培养用水,并加入少量微藻液,池中的休眠卵即可孵化,从而获得大量的轮虫。但原池保存休眠卵,由于病、敌害生物的危害,存活率不高。
★知识框架(必看)↓↓↓
★章节视频(必看)↓↓↓
★章节PPT↓↓↓
★章节文字稿↓↓↓
轮虫以休眠卵的方式进行保种,以渡过恶劣的外界环境。当环境适宜时,休眠卵将结束休眠,孵化出新的轮虫个体进行繁衍。请问休眠卵需要什么样的条件才会进行孵化呢?
已有的文献资料表明,休眠卵的孵化率与孵化时的环境状况、休眠卵形成时母体的生长状况、休眠卵的保存状况以及形成休眠卵的母体的遗传因素等有关。
下面我们将从休眠卵的孵化形式、休眠卵的孵化方法、影响休眠卵孵化因素三个方面学习轮虫休眠卵孵化的相关知识。
1、休眠卵的孵化形式:大多数种类轮虫的休眠卵孵化接具有两类形式。一类为零星孵化形式,意指轮虫的休眠卵在较长的一段时间内或多或少的以有规律的时间间隔而零星的孵化;另一类称为同步孵化形式,意指大量的休眠卵在休眠期后同时孵化。在水产养殖业的研究中,轮虫休眠卵的同步孵化至关重要。
2、休眠卵的孵化方法:可用各种玻璃培养缸、小水族箱为孵化容器,容器在使用前应清洗、消毒,再用过滤后的孵化用水冲洗干净,然后加入孵化用水。为了避免敌害生物的危害,清除小型甲壳动物,休眠卵孵化及培养仔虫的用水,需要用300目的筛绢网过滤。并调节水质至最适范围。
把少量轮虫休眠卵放入水中孵化。孵化容器放置在靠近窗口或有人工照明的位置。孵化期间每天需搅拌1~2次。如果条件适宜,一般在3~7天的时间内,即可孵化。
3、影响休眠卵孵化因素:主要从四个角度考虑:休眠卵保存的方法和时间对休眠卵孵化率的影响,孵化时的环境状况对休眠卵孵化率的影响,休眠卵形成时的环境状况对其孵化率的影响,品系或克隆间的差异。
首先,我们来学习休眠卵保存的方法和时间对休眠卵孵化率的影响。休眠卵保存的方法主要有原池保存、冷冻、干燥。根据相关的研究报道,将干燥后的休眠卵在48~61千帕大气压下进行听装密封,放于冷冻环境下保存,孵化率是最可靠的。可使休眠卵保存达6个月之久其孵化率也不会降低。
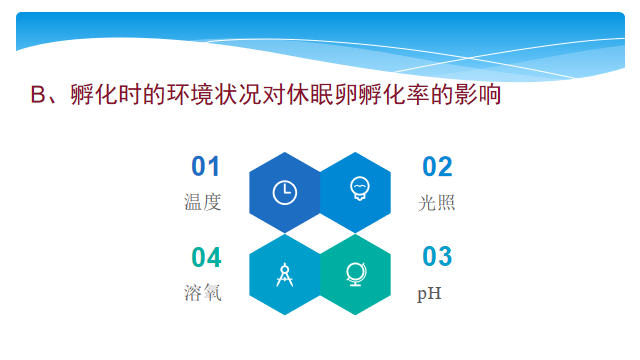
孵化时的环境状况对休眠卵的孵化率也会产生影响,其中主要的环境因素有:温度、光照、pH、溶氧。
温度对休眠卵孵化时间有明显的影响,休眠卵孵化的最适温度往往在该种群的适宜增长温度范围内。在孵化适宜温度范围内,孵化时间随温度的升高而缩短,二者呈现负相关关系。
在光照对轮虫休眠卵孵化率的影响常因种而异。如光对于红臂尾轮虫休眠卵、褶皱臂尾轮虫休眠卵的快速批量孵化是绝对必须的;对萼花臂尾轮虫休眠卵的孵化是有益的,但并非必不可少;但对角突臂尾轮虫和蒲达臂尾轮虫却无影响。然而有趣的是,光对角突臂尾轮虫新产休眠卵的孵化却是必要条件。产生上述差别的原因可能是由于新产休眠卵和经过一定时间保存的休眠卵对光的敏感性存在一定的差异造成的。
pH8时轮虫休眠卵孵化时间最短。有的种类休眠卵可孵化的pH范围较广,如萼花臂尾轮虫和角突臂尾轮虫休眠卵在pH4.5~11.5的范围内可以孵化。
萼花臂尾轮虫和角突臂尾轮虫休眠卵正常孵化的临界溶氧量为每升0.3㎎。
除以上四种环境因素外,最近的研究还表明,使用波长为320~380纳米的超声波对休眠卵进行辐射处理,可有效的提高休眠卵的孵化率。暴露于过氧化氢或前列腺素中,可使停滞发育的胚胎开始发育,即使在黑暗的环境条件下也一样。
在实际生产中,为了贮存休眠卵,把休眠卵置于休眠的状态是抑制其孵化的必要条件,贮存条件与促进休眠卵孵化的条件,恰恰是一对相矛盾的因子。如,在保存其休眠卵时:必需光照才能孵化的,其休眠卵需被置于暗处;喜暖性轮虫的休眠卵,则需要贮存在低温处;喜寒性轮虫的休眠卵置于高温下。
与之相反的,休眠卵孵化的环境条件与轮虫增长条件是相近的。如,休眠卵孵化的最适温度往往在该种群的适宜增长温度范围内。
孵化时的环境状况对休眠卵孵化率的影响的内容我们就讲解到这里。
接下来我们来讲解休眠卵形成时的环境状况对其孵化率的影响。休眠卵形成时,母体的生理状况、环境条件对轮虫休眠卵的孵化率具有较大的影响。
受精的混交雌体的摄食会影响其所产生休眠卵的孵化率,因为休眠卵形成时所需的能量远多于非需精卵,因此,母体的摄食对休眠卵的组成成分和生理状况有特别的影响。
比较了不同食物、温度和光照条件下形成的休眠卵的孵化过程,发现休眠卵形成时的环境状况并不能改变其孵化形式,但却能影响孵化时的环境因子与孵化率间的相互关系。通过改变休眠卵形成时的温度状况,可改变休眠卵孵化的最适温度,以及在该温度下的最大孵化率;以及改变休眠卵形成时培养液的盐度,也会影响其孵化时所需的最适盐度。
最后,研究发现,同种轮虫的不同克隆体所产的休眠卵不仅在形态上存在差异,其孵化形成也互不相同。对不同品系的褶皱臂尾轮虫在同样条件下形成和保存的休眠卵的孵化率的研究也表明,休眠卵的孵化率因品系的不同而异。


