第四节 影响群落组成和结构的因素
一、生物因素
群落结构总体上是对环境条件的生态适应,但在其形成过程中,生物因素起着重要作用,其中作用最大的是竞争与捕食。
(一)竟争对群落结构的影响
有不少研究实例说明竞争在群落结构形成中所起的重大作用。经典的是Lack 在 Galapagos 群岛上对达尔文莺(Darwin's finches)的研究。因竞争导致生态位的分化,而鸟类食性分化反映在鸟嘴形态上。直接的生态位分化的证据是,有的岛上只有一种在地面取食的鸟,其嘴长约10 mm,而在有两种或数种地面取食的鸟时,其最小型的嘴平均长8 mm,大一些的嘴平均长 12 mm,但没有10 mm 的。MacArhur 曾研究北美针叶林中林营(Dendroica)属的5种食虫小鸟,发现它们在树的不同部位取食,这是一种资源分隔现象,同样被解释为因竞争而产生的共存。
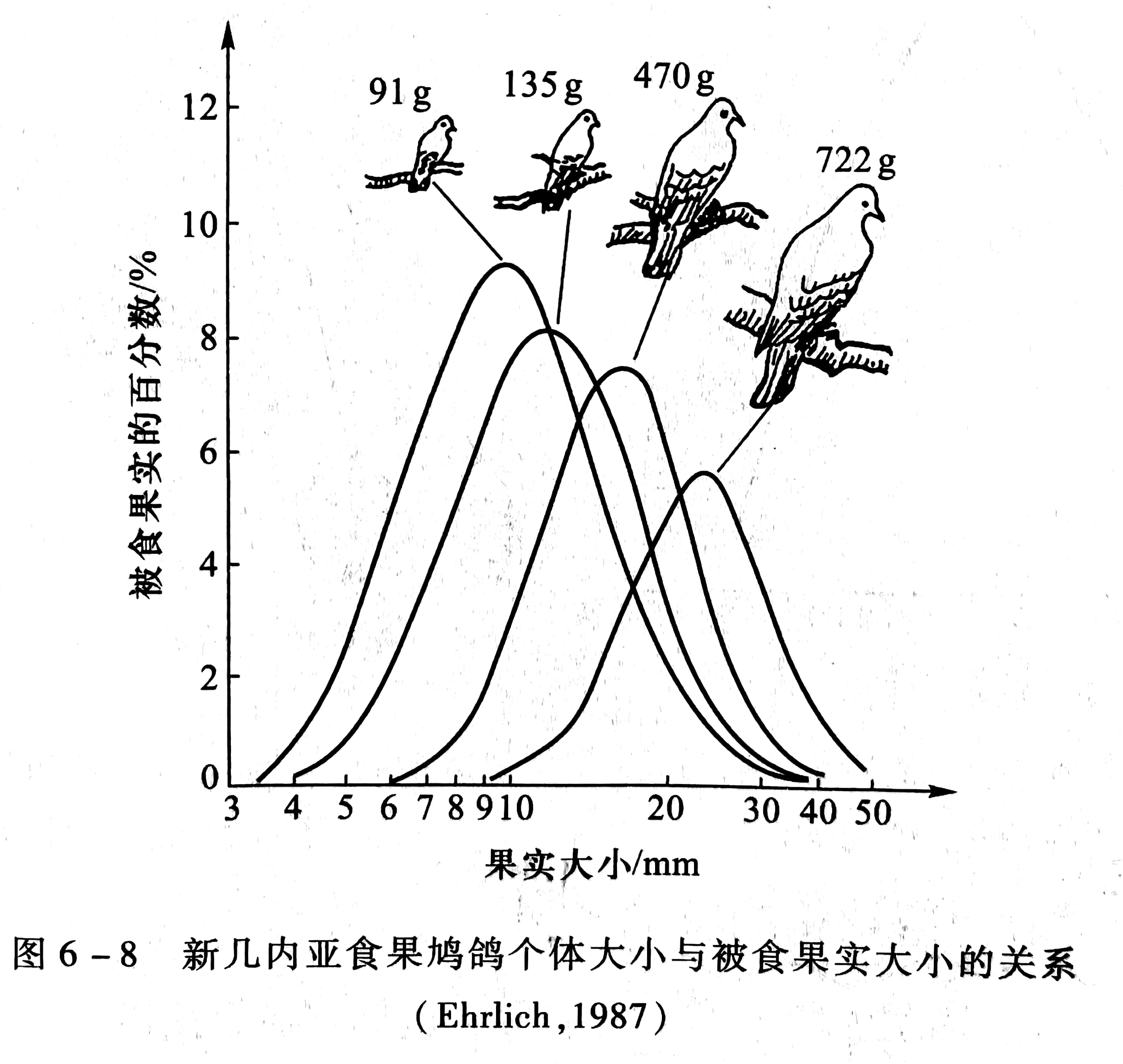
竞争和生态位分化研究还导致更广地应用个体大小特征作为资源分隔的指标。例如,在新几内亚有18 种食果鸠鸽,分属于果鸠(Ptilinopus )和皇鸠(Ducula)两属。在低雨林中,每一调查区最多8种,其体重大小的间隔大约是15 倍,最小的50 g,最大的800 go。个体大小影响取食果实大小的选择(图6-8)。
(二) 捕食对群落结构的影响
捕食对群落结构形成的作用,视捕食者是泛化种还是特化种而异。实验研究证明,随着泛化捕食者兔食草压力的加强,因兔把有竞争力的植物种吃掉,可以使竞争力弱的种生存,草地上的植物种数增加,所以多样性提高。但是吃食压力过高时,植物种数又随之降低,因为兔不得不吃适口性低的植物。因此,植物多样性与兔捕食强度的关系呈单峰曲线。
具选择性的捕食者对群落结构的影响与泛化捕食者不同。如果被选择的喜食种属于优势种,则捕食能提高多样性。例如潮间带常见的浜螺(Littorina litorea)是捕食者,吃很多藻类,尤其喜食小型绿藻(Enteromorpha),图6-9 表示随着浜螺捕食压力的增加,藻类种数也增加,捕食作用提高了物种多样性,其原因是藻类把竞争力强的浒苔的生物量大大压低了。但是,如果捕食者喜食的是竟争上占劣势的种类,则结果相反,捕食降低了多样性。
Paine(1966)在岩底潮间带群落中进行去除海星的试验。Paine 在一8 m长,2 m 宽的试验样地中连续数年把所有海星都去除掉,结果在几个月后,样地中藤壶成了优势种,以后藤壶又被贻贝所排挤,贻贝成为优势种。这个试验证明了顶级食肉动蜘成为决定群落结构的关键种(keystone species) ( 图6-10)。

二、干扰对群落结构的影响
干扰(disturbance)是自然界的普遍现象,就其字面含义而言,是指平静的中断,正常过程的打扰或妨碍。
生物群落不断经受着各种随机变化的事件,正如 Clements 指出的:“即使定最稳定的群丛也不完全处于平衡状态,凡是发生次生演替的地方都受到干扰的影响”。他们把干扰视为扰乱了顶极群落的稳定性,使演替离开了正常轨道。近代多数生态学家认为干扰是一种有意义的生态现象,它引起群落的非平衡特性,强调了干扰在群落结构形成和动态中的作用。
由大风、雪电、砍伐、火烧等引起的森林缺口;由放牧、动物挖掘、踩踏等造成的草原缺口,有的在干扰消失后会逐渐地恢复,但缺口也可能被周围群落的任何个种侵入和古有,并发展为优势者,哪一种是优胜者完全取决于随机因素,即先入侵的种取胜、至少在其一生之中为胜利者。当缺口的占领者死亡时,缺口再次成为空白,哪一种入侵和占有又是随机的。当群落由于各种原因不断地形成新的缺口,那么群落整体就有更多的物种可以共存,群落的多样性将明显提高,例如,澳大利亚的大堡礁,鱼类特别丰富,南部有 900 种,北部达1500 种,而珊瑚礁中每一直径3m左右的礁块中,可生活 50 种鱼以上。对如此高的鱼类多样性只以食物资源分隔是难以解释的,实际上许多鱼的食性是很接近的。
缺口形成的频率影响物种多样性,据此 Connell 等提出了中度干扰假说(intermediate disturbance hypothesis),即中等程度的干扰水平能维持高多样性。其理由是:(1)在一次干扰后少数先锋种入侵缺口,如某千扰频繁,则先锋种不能发展到演替中期,因而多样性较低;(2) 如果干扰间隔期很长,使演替过程能发展到顶极期,多样性也不很高;(3) 只有中等干扰程使多样性维持最高水平,允许更多的物种入侵和定居。
三、空间异质性与群落结构
群落的环境不是均匀一致的,空间异质性(spacial heterogeneity)的程度越高,意味着有更加多样的小生境,所以能允许更多的物种共存。
(一)非生物环境的空间异质性
Harman 研究了淡水软体动物与空间异质性的相关,他以水体底质的类型数作为空间异质性的指标,得到了正的相关关系:底质类型越多,淡水软体动物种数越多。植物群落研究资料说明,在土壤和地形变化频繁的地段,群落含有更多的植物种,而平坦同质土壤的群落多样性低。
(二) 生物空间异质性
R. H. MacArthur等曾研究鸟类多样性与植物的物种多样性和取食高度多样性之间的关系。取食高度多样性是对植物垂直分布中分层和均匀性的测度。层次多,各层次具更茂密的枝叶表示取食高度多样性高(结果是:鸟类多样性与植物种数的相关,不如与取食高度多样性相关紧密(图6-11)。对于鸟类生活,植被的分层结构比物种组成更为重要。因此,根据森林层次和各层枝叶茂盛度来预测鸟类,多样性是有可能的。在草地和灌从群落中,垂直结构对鸟类多样性就不如森林渡落重要,而水平结构,即镶嵌性或斑块性 patchiness)就可能起决定作用。

四、岛屿与群落结构
岛屿由于与大陆隔离,生物学家常把岛屿作为研究进化论和生态学问题的天然实验室或微宇宙。例如达尔文对 Galapagos 群岛的研究及 MacArthur 对岛屿生态学研究等。
(一)岛屿的种数 - 面积关系
岛屿中的物种数目与岛的面积有密切关系,通过许多研究得到证实,岛面积越大,种数越多(图6-12),并可以简单方程描述:
S=cAZ
或取对数
lgS =lgc + z(lgA)
其中:S——种数,A——面积,Z——种数-面积关系中对应的斜率,C——单位面积种数的常数。
Galapagos群岛面积与物种数的关系为S=28.6A0.32

广义而言,湖泊受陆地包围,也就是陆“海”中的岛,山的顶部成片岩石是低纬度中的岛,一类植被或土壤中的另一类土壤和植被斑块、封闭林冠中由于倒木形成的“林窗”(缺口),都可被视为“岛”。根据研究,这类“岛”中的种数 - 面积关系同样可以用上述方程进行描述。
(二) MacArthur 的平衡说
岛屿上的种数取决于物种迁入和灭亡的平衡;并且,这是一种动态平衡,不断地有物种灭亡,也不断地由同种或别种的迁入来补偿。平衡可用图6-13 说明。以迁入率曲线为例,当岛上无留居种时,任何迁入个体都是新的,因而迁入率高。随着留居种数加大,种的迁入率就下降。当种源库(即大陆上的种)所有的种在岛上都有时,迁入率为零。灭亡率则相反,留居种数越多,灭亡率越高。迁入率和灭亡率都取诀于岛的远近和大小,近而大的岛迁入率高,远而小的岛,则低。
迁入率曲线与灭亡率曲线交叉点上的种数,即为该岛上预测的物种数、根据平衡说,可说明下列四点:①岛屿上的物种数不随时间而变化;② 这是一种动态平衡,即灭亡种不断地被新迁入的种所代替;③大岛比小岛能“供养”更多的种;④ 随岛距大陆的距离由近到远,平衡点的种数逐渐降低。
(三)岛屿生态与自然保护
自然保护区在某种意义上讲是受其周围生境“海洋”所包围的岛屿,因此岛屿生态理论对自然保护区的设计具有指导意义。
一般说来,保护区面积越大,越能支持或“供养”更多的物种;面积小,支持的种数也少。但有两点需要说明:① 建立保护区意味着出现了边缘生境(如森林开发为农田后建立的森林保护区),适应于边缘生境的种类受到额外的支持:②对于某些种类而言,小保护区比大保护区可能生活得更好。
在同样面积下,一个大保护区好还是若干小保护区好,这决定于下列情况;①若每一小保护区内都是相同的一些种,那么大保护区能支持更多的种;② 从传播流行病而言,隔离的小保护区有更好的防止传播作用;③ 如果在一个相当异质的区域中建立保拉区、多不不保护区能提高空间的异质性,有利于保护物种多性性;④ 对密度低、增长率慢的大型动物,为了保护其遗传性,较大的保护区是必需的。保护区过小,种群数量过低,可能由于近交使遗传特征退化,也易于因遗传漂变而丢失优良物种的特征。
在各个小保护区之间的“通道”或走廊,对于保护是很有帮助的,它能减少被灭亡的风险,细长的保护区,有利于迁入。
五、平衡说和非平衡说
对群落结构形成的看法,有两种对立的观点,即平衡说和非平衡说。
平衡说认为共同生活在同一群落中的物种处于一种稳定状态。因为它们通过竞等、捕食和互利共生等种间相互作用而形成相互牵制的整体,在非干扰状态下群落的物种组成和数量都变化不大;群落实际上出现的变化是由环境的变化,即所谓的干扰引起的。总之,平衡说把生物群落视为不断变化着的物理环境中的稳定实体。
平衡说是由 C. Elton (1927)首先提出的,他认为群落中种群的数量是不断变化的,但其原因是由环境的变动,或由一个种群如被食者的种群变动导致捕食者种联变动。如果环境停止变动,群落将呈现稳定状态。R. H. MacArthur 主张动态平衡,在研究岛屿生物地理学中提出,群落的物种数是一常数,这是迁入和灭绝之间的平衡所取得的;因此构成群落的物种是在不断变化之中,而种数则保持稳定。
非平衡说认为,组成群落的物种始终处在不断地变化之中,自然界中的群落不存在全局稳定有的只是群落的抵抗性(群落抵抗外界干扰的能力)和恢复性(群落在受干扰后恢复到原来状态的能力)。非平衡说的重要依据就是中度干扰理论。Huston (1979)关于干扰对竞争结局的研究可以说明非平衡说。Lotka Volterra 的竞争排斥律可以被证明,但必须在稳急定而均匀的环境中,并且有足够的时间,才能使一种挤掉另一种,或通过生态位分化而共存。但在现实中环境是不断地变化的,种间竞争强度和条件有利于那一种都在变化之中,这可能是自然群落中竞争排斥证据有限的原因。
平衡说与非平衡说除对干扰的作用强调不同以外,一个基本区别是平衡说的注意焦点是系统处于平衡点时的性质,而对于时间和变异性注意不足;而非平衡说则把注意焦点放在离开平衡点时系统的行为变化过程,特别强调时间和变异性。平衡说认为,群落有向平衡点发展的趋势,但有或大或小的波动。因此,平衡说与非平衡说的区别在于干扰对群落重要作用认识上的不同。另一重要区别是把群落视为封闭系统还是开放系统。Lotka-Voltera 的竞争模型把两物种竞争视为封闭系统,结局是一种使另一种灭绝。开放系统的模型包括一组小室(模拟群落中的斑块性,斑块间可以有迁移存在),相互竞争中可能有一种灭绝,也可能由一小室迁入另一小室。模型研究证明:当系统被分为小室以后,由于小室之间高水平的连通性(connectedness),使达到平衡的时间大为延长。


