第五章 种内与种间关系
生物在自然界长期发育与进化的过程中,出现了以食物、资源和空间关系为主的种内与种间关系。我们把存在于各个生物种群内部的个体与个体之间的关系称为种内关系 (intraspeaific relationship),而将生活于同一生境中的所有不同物种之间的关系称为种间关系(Interspecific relationship)。随着科学技术的进步,这方面的研究亦越来越深入。天量的事实表明,生物的种内与种间关系除从光、温、水、及养料等方面去考虑竞争作用外,还包括有多种作用类型,是认识生物群落结构与功能的重要特性。
生物的种内关系包括有密度效应、动植物性行为(植物的性别系统和动物的婚配制度)、领域性和社会等级等。从生态学观点来讲,我们不能单从表面上和形式上看待种内关系。如森林中植物个体数量的增减,种内个体对矿质养分的需求和个体之间的遮阴关系,或动物中甚至同种内个体间为生存和争夺社会地位而进行的相互残杀等,从个体看,这种种内斗争是有害的,但对整个种群而言,因淘汰了较弱的个体,保存了较强的个体,从而有利于种群的进化与繁荣。因此,生物种内关系的研究,既应重视个体水平、亦应重视群体水平的研究。
从理论上讲,生物的种间关系有多种多样,但最主要的有9种相互作用类型(表5-1),可以概括为两大类,即正相互作用(positive interaction)与负相互作用( negative interaction)。

对于两个具体的物种而言,相互作用的类型可能会在不同的条件下有所变化,也可能在其生命史的不同阶段中有不同类型。例如,两个物种在某一时间可能是寄生作用,在另一时间则成为偏利作用,而在后来还可能是中性作用。因此,在种间关系的研究中,可利用野外调查和室内实验方法研究简化了的群落,将有助于区分各种相互作用类型和定量研究。
第一节 种内关系
一、密度效应
在种内关系方面,动物种群和植物种群的表现有很大区别,动物种群的种内关系主要表现为等级制、领域性、集群和分散等行为上;而植物种群则不同,除了有集群生长的特征外,更主要的是个体之间的密度效应(density effect),反映在个体产量和死亡率上。在一定时间内,当种群的个体数目增加时,就必定会出现邻接个体之间的相互影响,称为密度效应或邻接效应てthe effect of neighbours)。种群的密度效应是由矛盾着的两种相互作用决定的,即出生和死亡、迁入和迁出。凡影响出生率、死亡率和迁移的因子都起着调节作用,种群的密度效应实际上是种群适应这些因素综合作用的表现。根据影响因素的种类,可以将其作用类型划分为密度制约(density dependent)和非密度制约(density independent)。如可将气候因素、大气CO2,浓度等随机性因素看成是非密度制约因素,而将生物种间的捕食、寄生、食物、竞争等看成是密度制约因素。
根据生物种群密度效应的作用因素类型,可以将其划分为内源性与外源性作用因素。前者指内因,即种群自身内部的作用因素,它包括种内竞争所产生的各种作用因素,如遗传效应、病理效应和领域性效应等。后者指外因,即种群外部的作用因素,它包括种间竞争、食物和气候等外部作用因素所引起的密度效应。植物的密度效应已发现有两个基本的规律。
1. 最后产量恒值法则
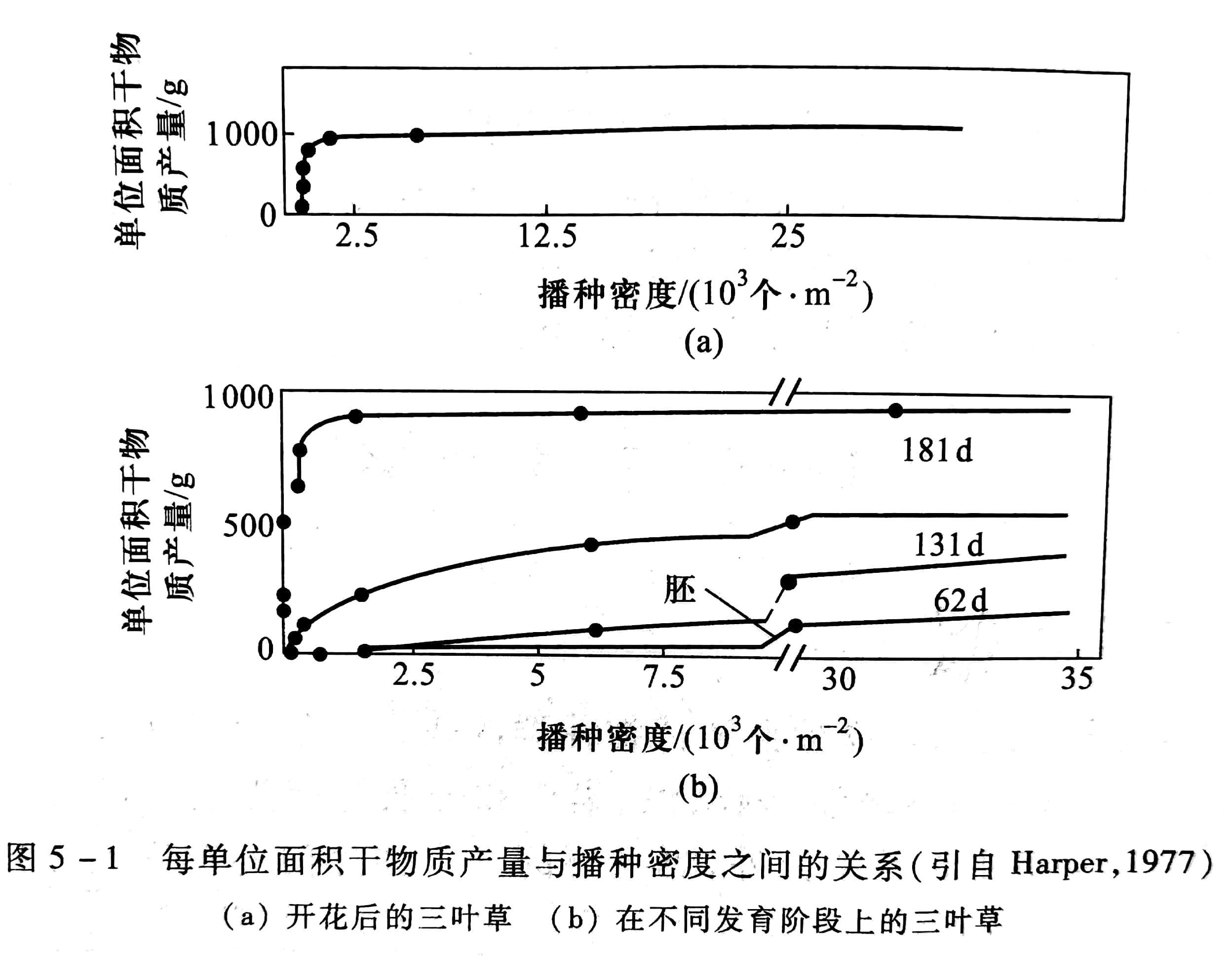
在一定范围内,当条件相同时,不管一个种群的密度如何,最后产量差不多总是一样的。图5-1 表示单位面积上三叶草的干物质产量与播种密度的关系:
(1) 开花后干物质产量随密度而变化,可以看出在密度很低时干物质随密度增加,但很快就趋于稳定。
(2) 在各种发育阶段上干物质随密度而变化,从萌芽初期到181 d,都呈现出产量最终恒定的规律。最后后产量恒值法则(the law of constant final yield)可用下式表示:
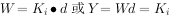
式中:W代表植物个体平均质量,d代表密度,Y代表单位面积产量,K, 为常数。最后产量恒值法则的原因在于:在高密度情况下,植株之间的光、水、营养物的竟争十分激烈,在有限的资源中,植株的生长率降低,个体变小。
2. -3/2 自疏法则
如果播种密度进一步提高和随着高密度播种下植株的继续生长,种内对资源的竞争不仅影响到植株生长发育的速度,进而影响到植株的存活率。在高密度的样方中,有些植株死亡了,于是种群开始出现“自疏现象”(self-thinning)。
Yoda 等(1963)把自疏过程中存活个体的平均株干重(W) 与种群密度(d)之间的关系用下式来表示:
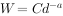
式中:a 是用密度/平均株干重的对数作图所得相关直线的斜率,C是该直线在纵坐标上(平均株干重的对数)的截距。
lgW = lgC – algd
Harper(1981)等对黑麦草(Lolium perenne)的研究发现a为 -3/2 区间内的一个恒值,因此 W=Cd"“这一经验公式被称为 -3/2 自疏法则(the -3/2 thinning law)。
在密度制约和非密度制约学说之间,还有一种将这两种理论综合起来的学说,即认为密度制约与非密度制约是同时起作用的。如 Kemp和 Keith(1970)在研究红松鼠种群与松树球果产量的关系中,证明食物决定了种群的数量,但气候控制了食物的量。
二、动植物的性行为
动、植物性行为的研究内容包括生物种群内部性别关系、动态以及决定性的环境因素。在营有性繁殖的种群内,种内相互作用首先表现在两性个体之间。种群的遗传特征及基因型多样性对于种群数量动态的重要意义使得动、植物性行为的研究在近年来受到了学者们的重视。
动、植物性行为与两个重要的生物学问题有关,即两性细胞的结合和亲代投入(parental investment)。亲代投入是指花费于生产后代和抚育后代的能量和物质资源。例如,有的动物产的卵大,有的卵小;有的一次生产的后代数很多,有的很少;有的精心抚育,有的置之不顾。这些都直接影响亲代投入的强度。
一般认为,有性繁殖是对生存在多变和易遭不测的环境下的一种适应性。因为雌雄两性配子的融合能产生更多变异类型的后代,在不良环境下至少能保证有少数个体生存下来,并获得繁殖后代的机会,所以多型性可能是一种很有效的对策。
(一)植物的性别系统
大多数植物种的个体具有雌雄两性花,是雌雄同花的。另一些植物种的个体具有雌雄两类花,雄花产生花粉,雌花产生胚珠,所以是雌雄同株而异花的,如玉米等。至于雌雄异株的植物,其雌花和雄花分别长在不同的植株上,只有这类一植物的雌性植株和雄性植株与动物中的雌体和雄体相当。此外,多年生草本三叶南星(Arisaema triphylum),在个体尚小时不开花,随着个体长大先有雄花,长到更大时才有雌花。当个体由大变小时,还可以出现相反的顺序。
植物界中,具雌雄异株的相当稀少,大约只占有花植物的5%左右,如银杏(Cinkgo biloba)等。雌雄异株多出现在热带具肉质果实的多年生植物中。多数生物学家认为,雌雄异株能减少同系交配的概率,具有异型杂交的优越性。此外,雌雄异株实际上是回避两性间竞争的对策,增加了两性利用不同资源的能力,也减少了食种子动物的压力。另一个环境压力由脊椎动物传粉造成,例如藤露兜树(Freycinetia reineckei),其植株多数是雌雄异株的, 只有含单性花的穗状花序,但偶然也出现雌雄同株的植株,具含雌雄两性花的花序。
(二)动物的婚配制度
1. 婚配制度的类型
婚配制度的类型按配偶数有单配偶制(monogamy)和多配偶制(polygamy),后者又分一雄多雌制(polygyny)和一雌多雄制(polyandry)。
(1) 一雄多雌制:一雄多雌制是最常见的婚配制度,例如海狗营集群生活,尤其在繁殖期内。通常雄兽首先到达繁殖基地,如岩岸、沙滩或大块浮冰,随种而异,争夺并保护领域。雌兽到达较晚,不久就生育幼仔。一只雄兽独占雌兽少则3只,多至40 只以上。
(2) 一雌多雄制:与一雄多雌制比较,动物中一雌多雄是很稀见的,据统计,鸟类中只占1%。典型的例子是距翅水雉(Jacana spinosa)。
(3)单配偶制:动物内单配偶制是比较少见的,只有鸟类以一雄一雌制较善遍,例如鸳鸯、天鹅、丹顶鹤和许多鸣禽。哺乳类中单配偶制倒是例外,如狐、鼬和河狸,其他脊椎动物各纲中单配偶制也有其例。
2. 影响婚配制度类型的环境因素
决定动物婚配制度的主要生态因素可能是资源的分布以及食物和营巢地在空间和时间上的分布情况。举例来说,如果有一种食虫鸟,占据一片具有高质食物(如昆虫)资源并分布均匀的栖息地,雄鸟在栖息地中各有其良好领域,那么雌鸟寻找没有配偶的雄鸟结成伴侣显然将比找已有配偶的雄鸟有利。这就是说,选择有利于形成一雄一雌制。不仅如此,如果雄鸟也参加抚育雏鸟,单配偶制也将比一雄多爾確制有利。因为在一雄多雌制中,雄性要抚育多个雌鸟所产的雏鸟,这几乎是不可能的。再者,在一个领域中巢窝数越少、个体密度越低,也将给雏鸟提供更多食物,降低受捕食者攻击的概率。由此可见,高质而分布均匀的资源有利于产生一雄一雌的单配偶制。
相反,如果高质资源是皇斑点状分布的,社群等级中处于高地位的雄鸟将选择并保卫资源最丰富的地方作为领域。在这种情况下,一旦占有资源丰富领域的雄鸟有了配偶以后,未有配偶的孤雌鸟选择配偶的困难将会增加。此时,一雄二雌的多配偶制就产生了。
(三)领域性
领域(territory)是指由个体、家庭或其他社群(social group )单位所占据的、并积极保卫不让回种其他成员侵人的空间。保卫领域的方式很多,如以鸣叫、气味标志或特异的姿势向人侵者宣告其领主的领域范围;以威胁或直接进攻驱赶人侵者等,称为领域行为0territorial behavior)。具领域性(territoriality )的种类在脊椎动物中最多,尤其是鸟兽,这些动物的高级神经活动最复杂。但某些节肢动物,特别是昆虫也具领域性。保护领域的目的主要是保证食物资源、营襄地、从而获得配偶和养育后代。在动物领域性的研究中,比较重要的概括性原理有:
(1)领域面积随领域占有者的体重而扩大(图5-2),领域大小必须以能保证供应足够的食物资源为前提,动物体重越大,需要资源越多,领域面积也就越大。
(2) 食物品质的影响,食肉性种类的领域面积较同样体重的食草性种类大,并且体重越大,这种差别也越大(图5-2的两条回归线斜率)。其原因是食肉动物获取食物更困难,需要消耗更多的能量,包括追击和捕杀。

(3) 领域行为和面积往往随生活史,尤其是繁殖节律而变化,例如鸟类一般在营巢期中领域行为表现最强烈,面积也大。
(四)社会等级
社会等级(social hierarchy)是指动物种群中各个动物的地位具有一定顺序的等级现象。等级形成的基础是支配行为,或称支配 - 从属(dominant-submissive )关系。例如:家鸡饲养者很熟悉鸡群中的彼此啄击现象,经过啄击形成等级,稳定下来后,低级的一般表示妥协和顺从,但有时也通过再次格斗而改变顺序等级。稳定的鸡群往往生长快,产蛋也多,其原因是不稳定鸡群中个体间经常的相互格斗要消耗许多能量,这是社会等级在进化选择中保留下来的合理性的解释。社会等级优越性还包括优势个体在食物、栖所、配偶选择中均有优先权,这样保证了种内强者首先获得交配和产生后代的机会。从物种种群整体而言,这有利于种族的保存和延续。社会等级制在动物界中相当普遍,包括许多鱼类、爬行类、鸟类和兽类。
社群等级和领域性这两类重要的社会性行为与种群调节有密切联系。Wyhne-Edwards 提出的种群行为调节学说的基础就是这种社会性行为与种群数量的关系。当动物数量上升很高时,全部最适的栖息地被优势个体占满。次适地段虽然成为其余个体的适宜栖息地,或者说可栖息密度具有一定弹性。随着密度增高,没有领域或没有配偶的从属个体比例也将增加,它们最易受不良天气和天敌的危害,这部分(称为剩余部分)比例的增加意味着种群死亡率上升,出生率下降,限制了种群的增长。相反,当种群密度下降时,这部分比例上升,种群死亡率降低,出生率上升,促进了种群的增长。


